Wiele aspektów stylu życia zwierzęcia związanych jest z okresem aktywności dobowej. Istnieją dwa główne wzorce aktywności: w ciągu dnia, kiedy zwierzę jest aktywne w ciągu dnia oraz w nocy, kiedy zwierzyna jest ekspansywna po zachodzie słońca w warunkach słabego oświetlenia. Niektóre zwierzęta mogą być żywotne głównie o świcie lub o zmierzchu, a jeszcze inne o każdej porze dnia (Hall, 2008). Wizualny system nieptasich teropodów jest przedmiotem stosunkowo niewielkiej ilości dyskusji, opartej głównie na ekstrapolacjach współczesnych zwierząt (Stevens, 2006). Najpopularniejszym założeniem stało się widzenie obuoczne u Tyrannosaurus rex (Stevens, 2006), jednak nakładające się pola widzenia w obu oczach niekoniecznie implikują stereopsję (Chure, 1998). Ogólna zasada obserwowana u współczesnych przedstawicieli teropodów – ptaków (Aves) jest taki, że model aktywności, którego używają znajduje odzwierciedlenie w budowie oka – patki nocne mają większą średnicę rogówki w stosunku do osiowej długości oka (prawdopodobnie jako adaptacja do zwiększenia wrażliwości wzrokowej na światło), natomiast ptaki dzienne mają większą osiową długość oka w stosunku do średnicy rogówki, a jej kształt koreluje z ostrością wzroku (Hall, 2008). Podobna zależność zaproponowano dla teropodów innych niż ptasie (Takanobu, 2011).
Różnice w formach oczu ptaków o różnych wzorcach aktywności: a). Siatkówka oczna, b). Soczewka oczna, c). grzebieniowa struktura naczyń krwionośnych należących do naczyniówki oka. Autor ilustracji Sibylle Schroer.
Wiele struktur kostnych koreluje z sąsiednia tkanką kostną. Narządy zmysłów w tym oczy mają mechaniczny wpływ na morfogenezę czaszki. Kiedy u zarodków kurzych upośledzone jest ‘’tworzenie się’’ oczu, anormalny jest również rozwój oczodołów w czaszce (Hall, 2008). Można się spodziewać, że rozmiar i kształt oka będą korelować z towarzyszącą mu anatomią kostną i można to wykorzystać do rekonstrukcji oczu niektórych wymarłych taksonów (Stevens, 2006). Sama morfologia czaszki niewiele mówi o układzie wzrokowym wymarłych teropodów, jednak oczodoły nieptasich teropodów wskazują uderzający zakres rozmiarów i kształtów, a różnorodność ta wydaje się mieć pewne znaczenie biologiczne i funkcjonalne. Do tej pory naczelne to jedyna grupa kręgowców dla której szczegółowo zbadano zależność między budową oka a orbitą. Wykazano, że u małych naczelnych gałka oczna wypełnia proporcjonalnie większą część orbity niż u większych (Schultz, 1940). Związek między wzorcami aktywności a anatomia kości badanych u naczelnych wykorzystano do interpretacji wzorców aktywności w taksonach kopalnych. Kay i Cartmill (1977) porównując tę cechę u ssaków tej samej wielkości stwierdzili, że szerokość otworu oczodołu w stosunku do długości czaszki pomaga odróżnić nocne i dobowe naczelne o długości czaszki mniejszej niż 70 mm. Kay i Kirk (2000) wykazali również, iż przy małych rozmiarach ciała naczelne nocne mają większą średnicę orbitalną niż naczelne dobowe. Heesy i Ross (2001) wykazali, że w przypadku osobników o długości czaszki mniejszej bądź równej 65 mm rozmiar orbity można oszacować wystarczająco dokładnie, aby ustalić charakter aktywności skamieniałych naczelnych, u których znana jest tylko 10 mm krawędź oczodołu. W przypadku naczelnych takich jak ssaki jedynym korelatem osteologicznym wielkość i kształt oka jest morfologia samej orbity. W przypadku braku samego oka można jedynie wywnioskować rozmiar i kształt tej jego części, która znajduje się bezpośrednio na orbicie i nie ma oczywistego sposobu oszacowania, która cześć gałki ocznej wystaje z kości. Ponadto, ponieważ objętość oczodołów naczelnych zwiększa się szybciej niż objętość oczu wraz ze wzrostem rozmiaru ciała, rozmiary oczodołów nie pozwalają dokładnie przewidzieć wielkości lub kształtu oczu przy zwiększonych rozmiarach ciała.
Jednak ptaki podobnie jak i większość gadów mają dodatkową strukturę kostną – pierścień twardówki, który niezawodnie koreluje z tkankami miękkimi oka. Pierścień twardówki to seria kosteczek połączonych w pierścień i umieszczonych w twardówce oka, przy czym wewnętrzna średnica pierścienia twardówki otacza średnicę rogówki. Dokładna struktura pierścienia twardówki różni się w zależności od gatunku gadów pod względem długości, stopniem skrzywienia, liczby i grubości kości. Sugerowano, że informacje o tkankach miękkich oka można wywnioskować z pomiarów pierścienia twardówki u ichtiozaurów, jaszczurek i ptaków (Hall,2008). Niestety w zapisie kopalnym rzadko są zachowane pierścienie twardówki co utrudnia rekonstrukcję oczu w wymarłych taksonach co istotnie koreluje z tkankami miękkimi oka.

Oczy kręgowców lądowych w tym ptaków (Hall,2008) są zwykle mniejsze niż oczodoły. Najbliższymi współczesnymi krewnymi teropodów (w tym ptaków) są krokodyle. Podczas gdy na przykład u koniowatych oczy zajmują ~75% wysokości i ~85% szerokości orbity, oczy Aligatora amerykańskiego (Alligator mississippiensis) mierzone po bokach na ~22 mm wypełniają tylko ~65% z 34mm orbity (Stevens,2006). Jednak oczy krokodyli nie mają pierścieni twardówkowych i mieszczą się w wyspecjalizowanych tkankach miękkich związanych z organizmami wodnymi, więc krokodyle prawdopodobnie nie są odpowiednim współczesnym odpowiednikiem co do potencjalnej rekonstrukcji oka teropoda (Stevens,2006). Prymitywne teropody takie jak: Coelophysis bauri, Eoraptor lunensis, Herrerasaurus ischigualastensis, Megapnosaurus rhodesiensis i Megapnosaurus kayentakatae mają duże zaokrąglone oczodoły. Cecha ta jest zachowana u wielu celurozaurów takich jak Ornitholestes, Compsognathus, Ornitomimów (Ornithomimidae), Owiraptorów (Oviraptoridae), Dromeozaurów (Dromaeosauridae), Terizinozaurów (Therizinosauria), Troodony (Troodontidae) i większości Tyranozaurów (Tyrannosauroidae – miedzy innymi: Albertosaurus libratus , Daspletosaurus torosus). Jest to prymitywny i najczęstszy ‘’stan’’ oczodołu u teropodów i wielu innych owodniowców. Jednak niezwykłe kształty oczodołów można znaleźć u teropodów z dużymi czaszkami, gdzie w swojej formie orbita jest prawie całkowicie podzielona na elementy grzbietowe i brzuszne. To zwężenie jest zwykle spowodowane przednią projekcja kości przedoczodołowej jak na przykład u : Abelisaurus comahuensis, Carcharodontosaurus saharicus, Carnotaurus sastrei i Tyrannosaurus rex. Stan ten jest do pewnego stopnia zmienny ontogenetycznie u tyranozaurów, niezwykłe zwężenie orbity występuje u holotypu Tarbosaurus baatar (PIN 551-1), podczas gdy czaszki mniejszych okazów (PIN 551-3 i 553-1) wykazują mniejszą separacje oczodołów przez kość oczodołową. W przypadku Acrocanthosaurus atokensis zwężenie jest spowodowane zarówno tylna projekcją kości łzowej jak i przednią projekcją kości zaoczodołowej . U teropodów które ‘’rozszczepiają’’ oczodół ( z wyjątkiem wspomnianego Tarbosaurus) część zawierająca oko jest grzbietowa i mniejsza co czyni je drapieżnikami o stosunkowo małych oczach. U Sinraptor dongi nieznacznie zachodzą na siebie zarówno kości łzowe jaki i podoczodołowe, ale orbita nie jest zwężona w takim samym stopniu jak u Acrocanthosaurus. Wiele teropodów o stosunkowo dość dużych łubach wykazuje stan pośredni między zaokrąglonymi i ściśniętymi kształtami orbitalnymi, gdzie najprostszym przykładem jest pionowo wydłużona orbita występująca u: Alioramus remotus , Ceratosaurus nasicornis , Torvosaurus tanneri i Yangchuanosaurus shangyuensis (u taksonów tych nie można dokładnie określić położenia oka, a zwłaszcza jego wielkości, aż do momentu potencjalnego odkrycia twardówki – Chure, 1998).
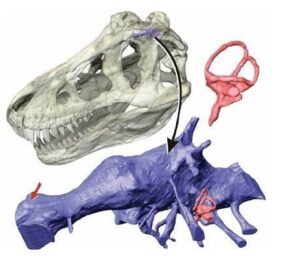
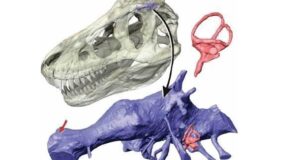
Chociaż sklerotyczne pierścienie u zdecydowanej większości teropodów nie są zbyt dobrze poznane , są jednak znane od gatunków Herrerasaurus ischigualastens, Megapnosaurus kayentakatae i Struthiomimus samueli, które wskazują, że u nieptasich teropodów o prostej (zaokrąglonej) strukturze orbity oko zajmowało prawie całą orbitę (Chure,1998) mniej więcej jak u współczesnych ptaków. Pierścienie twardówki znane są również od młodych tyranozaurów: Gorgosaurus libratus (Carpenter,2013) i Tarbosaurus baatar (Takanobu,2011). Zachowane pierścienie twardówki teropodów o bardziej złożonej strukturze orbitalnej pokazują jednak inny obraz. Zakładając, że rozmiar pierścienia twardówki łukuje izometrycznie z orbitą Carpenter (2013) obliczył rozmiar pierścienia twardówki Tyrannosaurus rex na podstawie pierścienia twardówki młodego Gorgosaurus. Co zaskakujące rozmiar orbity mniejszego Gorgosaurus był mniej więcej taki sam jak orbity Tyrannosaurus (okazu BHI 303 – ’’Stan’’), co sugeruje, że rozmiar oczu Tyrannosaurus nie zmieniał się znacząco wraz ze wzrostem rozmiaru czaszki.Szacunkowy pierścień twardówki BHI 3033 będzie miał około 7 cm i średnicę wewnętrznego otworu pierścienia twardówki około 3,5 cm. Na tej podstawie Stan z czaszką o długości ~140 cm od kości przedszczękowej do kwadratowo-jarzmowej ( Brunen, 2018) miał rogówkę o średnicy ~3,5 cm i źrenicę o średnicy ~2,5 cm. Średnica tęczówki oka u gadów jest zbliżona do średnicy pierścienia twardówki, a pełna średnica gałki ocznej (w zależności od głębokości oczodołu) jest około 1,4 – 1,5 razy większa od średnicy pierścienia twardówki. Zatem szacowana średnica gałki ocznej Stana wynosiła około 11 – 12 cm i około 13 cm w grzbietowo – brzusznym wymiarze orbity (Stolarz, 2013). Stevens (2006) uzyskał liczbę 91 – 119 mm na podstawie ekstrapolacji współczesnych zwierząt. Bliski krewny Tyrannosaurus rex, Tarbosaurus baatar jest potencjalnym wyjątkiem wśród dużych teropodów z grzbietową i brzuszną separacją oczodołów, z oczodołem grzbietowym zbliżonym do brzusznej średnicy (Chure, 1998). Przy długości czaszki 135,7 cm od kości przedszczękowej do kwadratowo-jarzmowej (Brunen, 2018) okaz Tarbosaurus baatar (PIN 551-1) średnica gałki ocznej ma około 13 cm długości w wymiarze grzbietowo – brzusznym co jest porównywalne do wielkości orbit BHI 3033 i wskazuje na porównywalny rozmiar oczu u obu zwierząt. Średnica pierścienia twardówki młodego Tarbosaurus baatar (MPC-D 107/7) z czaszką o długości ~29 cm od kości przedszczękowej do kwadratowo – jarzmowej zajmuje około 2/3 średnicy oczodołu. Co ciekawe u młodego osobnika nie podziału oczodołu na grzbietową i brzuszną, a duże wewnętrzne otwarcie pierścienia twardówki może wykazywać na dobrze rozwinięte widzenie w nocy (Takanobu, 2011).
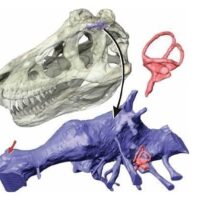
A. Czaszka młodego osobnika Gorgosaurus libratus z pierścieniami twardówki – B. Czaszka Tyrannosaurus rex (BHI 3033 ‘’Stan’’) z odtworzonymi pierścieniami twardówki według Carpentera (2013).
Chure (1998) podaje pomiary orbit i czaszki (od kłykcia przedszczękowego do kłykcia potylicznego) u wielu teropodów: Coelophysis bauri – długość czaszki od ~68 do 250 mm, długość oczodołu od ~20 do 40 mm, Compsognathus longipes – długość czaszki ~70 mm, długość oczodołu ~19 mm, Ornitholestes hermanni – długość czaszki ~138 mm, długość oczodołu ~35mm, Ceratosaurus nasicornis (USNM 4735) – długość czaszki ~660 mm, długość oczodołu ~77 mm, Tyrannosaurus rex (CMNH 7541) – długość czaszki ~572, długość oczodołu ~88 mm, Carnotaurus sastrei – długość czaszki ~596 mm, długość oczodołu ~80 mm, Monolophosaurus jiangi – długość czaszki ~670 mm, długość oczodołu ~85 mm, Allosaurus jimmadseni (MOR 693 ‘’Big Al.’’) – długość czaszki ~753 mm, długość oczodołu ~78 mm, Tyrannosaurus rex (AMNH 5027) – długość czaszki ~1355 mm, długość oczodołu 100 mm, Acrocanthosaurus atokensis (NCSM 14345 ‘’Fran’’) – długość czaszki ~1290 mm, długość oczodołu ~100 mm. Jak widać w powyższych danych od Chure proporcjonalnie do największe oczy miał Coelophysis bauri, gdzie oczodoły zajmowały około 29,4% długości czaszki, proporcjonalnie mniejsze oczy posiadały Acrocanthosaurus atokensis, którego oczodoły zajmowały ~7,5% długości czaszki oraz Tyrannosaurus rex z oczodołami zajmującymi ~8,3% długości czaszki (oczy Tyranozaurów są około 10-20% mniejsze niż część zajmowanych przez nie orbit). Należy jednak pamiętać, że oczy teropodów podobnie jak u ssaków zmniejszały się allometrycznie wraz ze wzrostem, dlatego proporcjonalnie mniejsze oczodoły niekoniecznie wskazują, że większe zwierzę jest mniej skupione na wzroku. Może to być związane ze zmianami ontogenetycznymi u dużych teropodów takich jak Tyrannosaurus rex i Tarbosaurus baatar (Chure,1998 – Takanobu,2011 – Carpenter,2013).
Oczodoły poszczególnych teropodów według Chure (1998): A. Eoraptor lunesis – B. Tyrannosaurus rex (CMNH 7541) – C. Ceratosaurus nasicornis – D. Torvosaurus tanneri – E. Yangchuanosaurus shangyuensis – F. Sinraptor dongi – G. Allosaurus jimmadseni (MOR 693 ‘’Big Al.’’) – H. Monolophosaurus jiangi – I. Cryolophosaurus ellioti – J. Carcharodontosaurus saharicus – K. Tyrannosaurus rex – L. Acrocanthosaurus atokensis – M. Carnotaurus sastrei – N. Abelisaurus comahuensis
*Opublikowany artykuł może być wykorzystywany wyłącznie na własny użytek edukacyjny, nie może stanowić źródła informacyjnego dla innych blogów, wykluczonym jest także udostępnianie ani publikowanie go bez zgody autora.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Poprzedni artykuł:
Dziesięć największych osiągnięć paleontologii związanych z dinozaurami ostatniej dekady Następny artykuł:
Mary Anning
Dziesięć największych osiągnięć paleontologii związanych z dinozaurami ostatniej dekady Następny artykuł:
Mary Anning