Jednym z najbardziej charakterystycznych jurajskich drapieżnych dinozaurów był bez wątpienia dość duży, a według niektórych przesłanek gigantyczny i licznie występujący rodzaj – Allosaurus, zamieszkujący zachodnią Amerykę Północną i prawdopodobnie zachodnie regiony współczesnej Europy pod koniec okresu jurajskiego ery mezolitu (~157 do ~145 mln lat temu). Kiedy został formalnie opisany i ‘’wprowadzony’’ do świata naukowego przez słynnego paleontologa Othniela Charlesa Marsha w 1877 roku (niemalże trzy dekady przed opisem Tyrannosaurus rex) był największym, nieco lepiej znanym drapieżnym teropodem. W związku z tym zaczęto Allosaurus uważać za niebezpiecznego drapieżnika zdolnego do łowów i uśmiercenia niemal każdej zdobyczy począwszy od małych ornitopodów po gigantyczne zauropody (zwłaszcza jeżeli polował w stadach). Ślady na kościach roślinożernych dinozaurów oraz uszkodzone kręgi samego Allosaurus dowodzą, że aktywnie atakowały i zabijały swoją zdobycz, choć prawdopodobnie często ceną natarcia były liczne urazy a nawet utrata życia. Nowo opublikowana praca naukowa Camerona C.Pahla oraz Luisa A.Ruedasa przedstawia nieco inny pogląd na tego drapieżnika ekosystemów późnej jury.

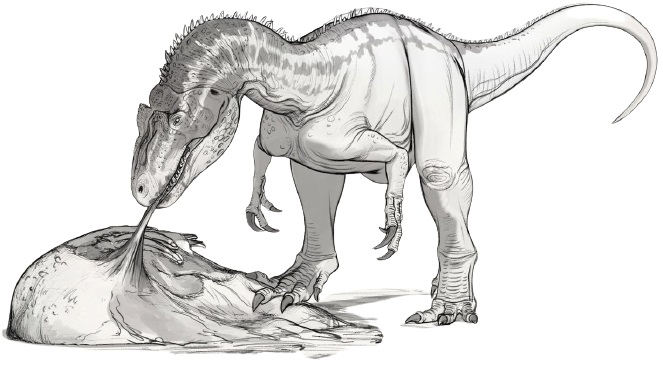
Model przedstawiający potencjalny wygląd anatomiczny oraz umaszczenie jednego z dwóch obecnie ważnych gatunków – Allosaurus jimmadseni, który był gatunkiem poprzedzającym w czasie Allosaurus fragilis i najprawdopodobniej jego bezpośrednim przodkiem rozwojowym. Ogólnie Allosaurus jimmadseni jako gatunek cechował się nieco węższą i mniej masywną czaszką niż jego młodszy krewny Allosaurus fragilis, możliwe więc, iż drapieżnik ten skupiał się na polowaniu na mniejsze i łatwiejsze ofiary, być może specjalizował się bardziej w łowach na ornitopody, heterodontozaury oraz mniejsze teropody. Autor: Li Ke Mnaga.
W zeszłym roku opublikowano badanie analizujące 2368 skamieniałości odkrytych od 1981 roku na stanowisku Mygatt-Moore Quarry w zachodnim Kolorado ukazujące, że 29 procent odkrytego materiału zawierało ślady zębów dużych teropodów, w tym w bardzo dużej mierze allozaurów. Paleontolodzy ujawniają, iż ciałami martwych dinozaurów żywiły się całe multum allozaurów i nie były to tylko truchła stegozaurów i zauropodów ale także innych allozaurów, co mogło by potwierdzać okazjonalny kanibalizm. Olbrzymie nagromadzenie kości dinozaurów w tej części słynnej formacji Morrison jest tłumaczone przez badaczy jako rodzaj naturalnej pułapki. Chociaż liczba zadrapań odpowiadająca zębom allozaurów jest niezwykle wysoka i nie odpowiada podobnym statystyką z innych lokacji, jest prawie pewne, że allozaury były bardzo chętne ku padlinożerności. Nowa publikacja wysuwa jeszcze dalej idące wnioski, z których wynika, że allozaury były fakt faktem drapieżnikami, co najlepszymi padlinożercami. W rzeczywistości autorzy porównują je do odpowiedników dzisiejszych sępów i kondorów.
Para naukowców z Portland State University wysunęła na swój sposób odważne stwierdzenie poprzez symulacje ekologiczne pokazujące, że allozaury nie musiały przede wszystkim polować, aby przeżyć, gdyż mogły uzyskać wystarczającą ilość kalorii z ogromnych tusz zauropodów i innych dinozaurów, które były dość powszechne w późnej jurze (allozaury koegzystowały wraz z gigantycznymi zauropodami takimi jak: Apatosaurus, Diplodocus, Brachiosaurus, Supersaurus czy Maraapunisaurus). Modelowanie ówczesnych ekosystemów wykazało, że tusze zauropodów statystycznie wystarczały do wykarmienia całej populacji olbrzymich drapieżników, które nie musiały nawet zbyt często aktywnie polować. Autorzy pracy naturalnie poparli swoje wnioski innymi badaniami, zbadali między innymi potencjalne wymagania żywieniowe allozaurów i średnie występowanie ‘’zwłok’’ o określonej wartości odżywczej, relacje ekologiczne i sieci troficzne zwierząt w formacji Morrison a także cechy morfologiczne allozaurów.
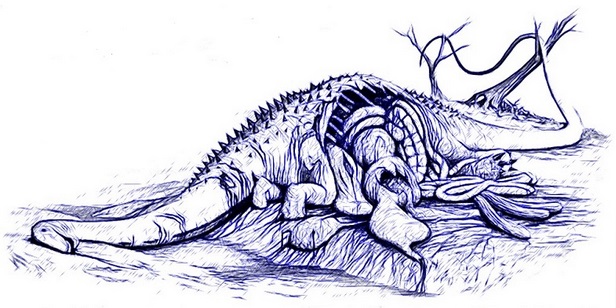
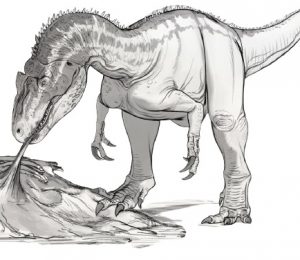
Odkrycia z niektórych miejscach sugerują, iż dziesiątki allozaurów i współistniejących z nimi innych drapieżnych dinozaurów uczestniczyły stosunkowo często w padlinożernych ‘’orgiach’’, które zapewniały im zwłoki roślinożernych gigantów, tak że nie musiały one zbyt często aktywnie polować. Autor szkicu: Bob Nicholls.
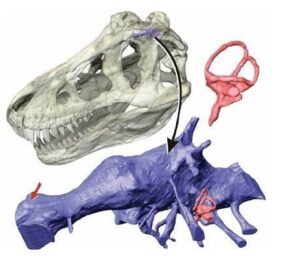
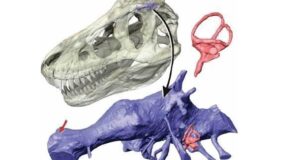
Niektóre wcześniejsze badania anatomii allozaurów pokazują, że ten teropod miał względnie słaby chwyt szczękowy, a jego czaszka i zęby były stosunkowo kruche (bardziej nakierunkowane na ‘’krojenie’’ i gryzienia żywej i martwej zwierzyny w miękkie części ciała niż do miażdżących ugryzień) i nie nadawały się do aktywnego drapieżnictwa. Ponadto stopień widzenia obuocznego u tego drapieżnika wynosił tylko 30 procent w stosunku do Tyrannosaurus rex i 15 procent w parowaniu z dzisiejszym Panthera leo. Na podstawie tych wszystkich danych autorzy pracy wywnioskowali, że allozaury były ‘’dominującymi padlinożercami’’ w ekosystemach formacji Morrison i prawdopodobnie wszystkich innych, w których występowały, a zatem nie reprezentowały domniemanych drapieżników wierzchołkowych. Z tych wszystkich danych nasuwał by się niepodważalny wniosek, że allozaury bytujące w środowisku obfitującym w tysiące ton zdechłych tusz musiały rozwinąć się jako specjaliści w pochłanianiu trucheł. Ale czy to prawda?, czy allozaury były naprawdę pasywnymi łowcami, którzy ograniczali się do polowania bardzo rzadko?, skoro nawet dzisiejsi padlinożercy takie jak choćby hieny od czasu do czasu polują bardzo umiejętnie.
Ponadto znaleziska skamieniałości w postaci patologii szkieletowych pokazują, że allozaury konfrontowały się z wciąż żywymi i aktywnie broniącymi się przed nimi roślinożercami zdaję się to potwierdzać choćby materiał dorosłego osobnika odkrytego w osadach formacji Morrison, który ma zdeformowaną kość łonową, gdzie uraz tkanki kostnej ma objętość pomarańczy, a kształt i wielkość rany (otwór stożkowy) z bardzo dużym prawdopodobieństwem odpowiada kolcom ogonowym Stegosaurus. Uraz doprowadził do wdania się infekcji, która rozprzestrzeniła się na tkanki (jelita i narządy rozrodcze zwierzęcia), a biorąc pod uwagę brak oznak gojenia się kości wydaje się prawdopodobne, iż okaleczenie było śmiertelne.
Z wnioskiem z tego eseju można się częściowo zgodzić, że niewątpliwie wiele licznych allozaurów wykorzystywało zwłoki dziesiątek ton ległych zauropodów jako ważne źródło ich składników odżywczo pokarmowych, wyjaśnia on także kwestię tak dużego nagromadzenia kości allozaurów i innych teropodów w niektórych lokajach. Jednak jedynym punktem, z którym bym polemizował z autorami była by potencjalna wyłączna padlinożerność i stosunkowo rzadka chęć do polowania na żywą zdobycz.

Statuetka odzwierciedlająca scenę agresywnej interakcji pomiędzy Allosaurus fragilis a Stegosaurus stenops. Jak udowadniają nam niektóre z odkryć paleontologicznych rosłe Stegosaurus z pewnością nie były łatwą zwierzyną łowną, a drapieżnik nie zawsze musiał być zwycięzcą gwałtownej konfrontacji, która mogła zakończyć się dla niego rozległymi urazami a w najgorszym wypadku nawet ‘’śmiercią’’. Dlatego najbardziej prawdopodobnym jest fakt, iż atakowały je tylko bardzo głodne i zdesperowane teropody, które nie mogły upolować łatwiejszej zdobyczy bądź alternatywnie znaleźć zasobów pokarmowych od dłuższego czasu. Autor: statuetki Joe Wang.
Bibliografia użyta w realizacji tej odsłony artykułu:
Chure, D. J. (2000). Prey bone utilization by predatory dinosaurs in the Late Jurassic of North America, with comments on prey bone use by dinosaurs throughout the Mesozoic (PDF). Gaia. 15: 227–232. ISSN 0871-5424
Loewen, M. A.; Sampson, S. D.; Carrano, M. T.; Chure, D. J. (2003). Morphology, taxonomy, and stratigraphy of Allosaurus from the Upper Jurassic Morrison Formation. Journal of Vertebrate Paleontology. 23 (3): 72A. doi: 10.1080/02724634.2003.10010538
Anton, M.; Sánchez, I.; Salesa, Manuel; Turner, A. (2003). The muscle-powered bite of Allosaurus (Dinosauria; Theropoda): An interpretation of cranio-dental morphology (PDF). Estudios Geologicos. 59 (5): 313–323. doi: 10.3989/egeol.03595-6106
Bakker, R. T.; Bar, G. (2004). Dinosaur crime scene investigations: theropod behavior at Como Bluff, Wyoming, and the evolution of birdness. In Currie, Philip J.; Koppelhus, Eva B.; Shugar, Martin A.; Wright, Joanna L. (eds.). Feathered Dragons: Studies on the Transition from Dinosaurs to Birds. Bloomington and Indianapolis: Indiana University Press. (str. 301–342). ISBN 978-0-253-34373-4
Carpenter, K.; Sanders, F.; McWhinney, L. A.; Wood, L. (2005). Evidence for predator-prey relationships: Examples for Allosaurus and Stegosaurus. In Carpenter, Kenneth (ed.). The Carnivorous Dinosaurs. Bloomington and Indianapolis: Indiana University Press. (str. 325–350). ISBN 978-0-253-34539-4
Stevens, K. A. (2006). Binocular vision in theropod dinosaurs. Journal of Vertebrate Paleontology. 26 (2): 321–330. doi: 10.1671/0272-4634(2006)26[321: BVITD]2.0.CO;2
Snively, E.; Cotton, J. R.; Ridgely, R.; Witmer, L. M. (2013). Multibody dynamics model of head and neck function in Allosaurus (Dinosauria, Theropoda). Palaeontologia Electronica. 16 (2). doi: 10.26879/338
Drumheller, S. K.; et al. (2020). High frequencies of theropod bite marks provide evidence for feeding, scavenging, and possible cannibalism in a stressed Late Jurassic ecosystem. PLOS ONE. 15 (5): e0233115. doi: 10.1371/journal.pone.0233115
Pahl, C. C.; Ruedas, L. A. (2021). Carnosaurs as apex scavengers: Agent-based simulations reveal possible vulture analogues in Late Jurassic dinosaurs. Ecological Modelling. 458: 109706; doi: 10.1016/j.ecolmodel.2021.109706
Treść merytoryczna opublikowanego artykułu na łamach witryny DinoAnimals.pl może być wykorzystywana jedynie na własny użytek edukacyjny i nie może stanowić podmiotu informacyjnego dla innych blogów. Wykluczonym jest także udostępnianie ani publikowanie go poprzez kanały tematyczne bez zgody jego autora.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Poprzedni artykuł:
Niszczyciel goleni z późnej kredy Następny artykuł:
Dinozaur z keratynowymi „wstążkami”
Niszczyciel goleni z późnej kredy Następny artykuł:
Dinozaur z keratynowymi „wstążkami”