
W dziejach naszej planety latające zwierzęta wyewoluowały wielokrotnie szerokie zakresy rozmiarów skrzydeł. Wśród nich znalazły się wyjątkowo duże gatunki ptaków jak Pelagornis sandersi (z późnego oligocenu, circa 28 – 25,72 mln lat temu), którego maksymalną rozpiętości skrzydeł uwzględniając pióra szacuje się na ~7,36 metra i wagę ~41 kilogramów czy Argentavis magnificen (z górnego miocenu, około 8 –6,3 mln lat temu) o szczytowym ~7 metrowym zakresie narządu lotu i masie ciała w zakresie ~59 kilogramów. Kilka dużych gatunków pterozaurów pojawiło się w okresie kredowym. Precyzuje się, iż skala skrzydeł Tropeognathus mesembrinus (największego pterozaura inadekwatnego od familii Azhdarchidae z wczesnej kredy w przybliżeniu 113 –110 mln lat temu) oscylowała w zakresie 8,26 metra, a gramatura na ~66 kilogramów, natomiast u Geosternbergia sternbergi (występujący w późnej kredzie ~81,5 –80,5 mln lat temu) miara płatów nośnych orientacyjnie wynosiła ~7,25 metra, a ciężar na ~36 kilogramów. Mimo to rozmiary wcześniej wymienionych są deklasowane przez pewnych przedstawicieli jednej z najbardziej udanych grup pterozaurów – Azhdarchidae (dobrze reprezentowane na lądach Gondwanu i Laurazji) obejmujących kilka bardzo dużych gatunków uważanych za jedne z największych stworzeń skłonnych do potencjalnego wznoszenia się w powietrze w historii globu wśród których sytuują się Quetzalcoatlus northropi z Mastrychtu w Stanach Zjednoczonych, Arambourgiania philadelphiae z Mastrychtu w Jordanii, Cryodrakon boreas z Kampanu w Kanadzie, Thanatosdrakon amaru z Santonu w Argentynie oraz podmiot prezentowanej przeze mnie poniższej publikacji Hatzegopteryx thambema z Mastrychtu w Rumunii.
Autorzy opisowi: Éric Buffetaut, Dan Grigorescu, Zoltán Csiki-Sava (2002).
Etymologia: Nazwa rodzajowa Hatzegopteryx jest kombinacją rumuńskiego słowa Hatzeg, która nawiązuje do dorzecza rzeki w mieście Hatzeg zlokalizowanego w Rumunii oraz staro greckiego pteryx oznaczającego skrzydło. Epitet gatunkowy thambema również wywodzi się ze antycznej greki i znamieniowi potwora, nawiązując do rozmiarów zwierzęcia.
Wymowa fonetyczna: Hat – zeh – gop – teh – rix.
Dystrybucja geograficzna: Europa rejony współczesnej Rumunii – formacje Densuș-Ciula, Sânpetru oraz Sebeş.
Datowanie: Późna kreda (Mastrycht) około 71 – 68 mln lat temu.
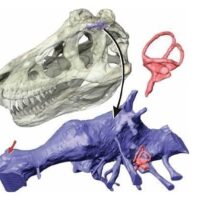
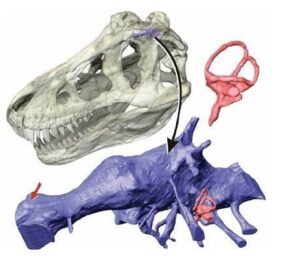
Szacunki gabarytowe: Holotyp tego azdarcha określony numerem katalogowym FGGUB R 1083A szacowany jest na ~314 kilogramów masy (tegoroczna kalkulacja dokonana przez Petera Menszikova, gdzie do obliczenia tonażu wykorzystany został model 3d wykonany za pośrednictwem programu Pixologic ZBrush i poddany wydrukowi fizycznemu przy użyciu drukarki Creality K1, z kolei rachuba wolumetryczna przeskalowanych poszczególnych wzorców elementów ciała przeprowadzona została przy użyciu aplikacji Surface Volume 3D Analist, kierując się wzorem m = ρ * V, gdzie m definiuje masę, ρ oznacza gęstość natomiast V symbolizuje objętość ), ~9,37 metrową rozpiętość skrzydeł oraz ~3,98 metra wysokości w naturalnej pozycji lokomocyjnej od styku kończyn z podłożem do szczytu łba.
Reprezentacja zachowanego materiału skalnego: Holotyp oznaczony jako FGGUB R 1083A personifikują dwa fragmenty tyłu czaszki (obszar potyliczny i podniebienny) oraz niekompletnej lewej kości ramiennej.
· FGGUB R1625 – środkowa część kości udowej o długości ~38,5 cm (kompletna kość nobilitowała w granicy 40 centymetrów) wydobyta w pobliżu Rumuńskiej miejscowości Tuştea.
· EME 315 – siódmy kręg szyjny (Hatzegopteryx sp.) o około 24 centymetrowej długości (cały kręg pierwotnie oscylował w granicy 30 centymetrów) pozyskany na stanowisku Râpa Roşie.
· EME 215 – duży kręg szyjny odnaleziony na obszarze Râpa Roșie (formacja Sebeș ).
· LPB (FGGUB) R.2347 – Częściowo zachowana obecnie największa rozmiarowo przednia część żuchwy potencjalnie przypisywana do Hatzegopteryx sp. odkryta w strefie Rumuńskiej wsi Vălioara w okręgu Hunedoara w gminie Răchitova, a zlokalizowana w zbiorach paleontologicznych Laboratorium Paleontologii Wydziału Geologii i Geofizyki Uniwersytetu w Bukareszcie.
Pozyskany został także nieskatalogowany materiał przydzielany do Hatzegopteryx sp. w postaci kości łopatkowo – kruczej, częściowo zachowanej obręczy barkowej, kawałków kręgów szyjnych, paliczka ‘’śródręcza’’ , fragmentu kość obręczy barkowej oraz fragment kości ramiennej z miejscowości Vadu (gmina Sântămăria-Orlea) – formacja Sânpetru.
Systematyka: Pterosauria · Zambellisauria · Macronychoptera · Novialoidea · Breviquartossa · Pterodactylomorpha · Monofenestrata · Pterodactyliformes · Caelidracones · Pterodactyloidea · Lophocratia · Eupterodactyloidea · Ornithocheiroidea · Azhdarchoidea · Neoazhdarchia · Neopterodactyloidea · Azhdarchidae · Quetzalcoatlinae.

Teraźniejsza rekonstrukcja potencjalnego kształtu anatomicznego i szaty pigmentacyjnej osobnika płci męskiej gatunku Hatzegopteryx thambema, jednego z największych kręgowców podatnych do implicytnycznego lotu w historii naszej planety. Kreator statuetki i wersji ubarwienia: Cen Dao Yi.
Deskrypcja informacyjna:
· Z relatywnie dużą dozą prawdopodobieństwa inicjalne szczątki Hatzegopteryx thambema mogły zostać zidentyfikowane już w 1899 roku przez węgierskiego arystokratę Franza Nopcsa von Felső-Szilvás uważanego powszechnie za jednego z twórców paleobiologii. Natomiast w 1991 roku, gdy po raz pierwszy zaprezentowano materiał skalny tego azdarcha środowisku paleontologicznemu, gdzie ze względu na rozmiar fragmentów szkieletu nie dawano pierwotnie wiary w to, iż może on należeć do pterozaura. Badacze (między innymi David B. Weishampel i współautorzy) przypisali, więc skamieniałości do niezidentyfikowanego jeszcze teropoda o ‘’silnych’’ proporcjach ciała (błąd był łatwy do popełnienia ze względu na znacznie większą powszechność dużych teropodów niż dużych pterozaurów). Dopiero w 2002 roku francusko – rumuński zespół badaczy (wymienionych powyżej) ostatecznie ustalił, że ‘’surowiec’’ szkieletowy należy do nieznanego wcześniej nauce pterozaura. Niestety i w tym przypadku dać o sobie znała ‘’klątwa gigantycznych pterozaurów’’ objawiająca się bardzo skromną liczbą zachowywanych fragmentów kopalnych, co znalazło odzwierciedlenie także u rodzaju Hatzegopteryx. Mimo wszystko znane elementy są na tyle duże by sugerować, że ich pierwotny ‘’właściciel’’ osiągał monumentalne rozmiary jak na przedstawiciela pterozaurów. Ponadto fosylia podczas analizy wykazały oznaki wewnętrznego stwardnienia w postaci grubszych ścian kości (4 – 6 mm) co jest differentią nawet dwukrotnie większą niż tą obserwowaną u większości innych wielkich przedstawicieli rodziny Azdarchidae (dla przykładu u Arambourgiania philadelphiae wartość ta wynosi ~2,6 mm).
· Hatzegopteryx bytował na wyspie Hateg (terytorium współczesnej Rumunii) dużej i dawno nieegzystującej wyspy na morzu Tetydy oddzielonej o co najmniej 200 kilometrów od najbliższego pasma lądowego, której najbardziej precyzyjne szacunki powierzchni w etapie genologicznym Mastrychtu (~72,1 – 66 mln lat temu) wynoszą ~80.000 tysięcy kilometrów kwadratowych (co odpowiada mniej więcej rozmiarowi obszarowemu współczesnej Nowej Gwinei czy Hokkaido). Wyspa powstała w wyniku wypiętrzania tektonicznego podczas początkowej orogenezy alpejskiej, na skutek ruchów płyty Adriatyku przez tak zwany Ocean Piemontsko – Liguryjski, a jej położenie w tym czasie odpowiadało pasowi równikowemu (około 27° północnej szerokości geograficznej). Analizy paleoklimatyczne ujawniły, że klimatyp na Hatzeg był subtropikalny (z wyraźnym podziałem na pory deszczowe i suche) ze średnimi rocznymi temperaturami pomiędzy 20° C – 25° C, gdzie jej środowisko zdominowane było przez pasma fluwialno- jeziorne oraz w przeważającym stopniu otwarte pasma lasów iglastych i w nieco mniejszym liściastych.
· Interesującym faktem jest również występująca na wyspie endemiczna fauna osiągająca niższe wartości długościowo wagowe od swoich kontynentalnych krewnych – zwłaszcza niepokaźne formy zauropodów (Magyarosaurus dacus czy Paludititan nalatzensis). Zjawisko to jest efektem tak zwanej karłowatości wyspiarskiej (po raz pierwszy przedstawiona przez wcześniej wspomnianego Franza Nopcsę w Wiedniu 1912 roku, który znacząco przyczynił się do poznania szaty zwierzęcej Hateg) będącej selektywnym procesem zmniejszania gabarytów ze względu na odizolowanie od zewnętrznych zasobów pokarmowych oraz ograniczenia zasięgu populacyjnego do mniejszego środowiska rezydowania, takiego jak wyspy. W ekosferze pozbawionej obecności wielkich teropodów (największego z nich reprezentował Balaur bondoc z rozmiarami ~2,2 metra długości i masie ~31 kilogramów) Hatzegopteryx ze swoją kubaturą personifikował drapieżnika szczytowego łańcucha pokarmowego (jest to jedyny przypadek w dziejach ery mezozoicznej, gdzie pterozaur zajmował miejsce łowcy krańcowego).
· Wątek predyspozycji do latania Hatzegopteryx i innych zogromniałych przedstawicieli Azdarchidae jest tematyką niezliczonych debat. Środowisko paleontologiczne, półprofesjonalne oraz niespecjalistyczne jest podzielone między opinią, że były to zwierzęta zbyt pokaźne masowo, aby mogły zainicjować start z miejsca i opinią, iż były w stanie wystartować używając tak zwanego ‘’wystrzelenia czworonożnego’’ – skaczącą sekwencje inauguracyjną, aby wejść w lot zwłaszcza podczas startu z ziemi. W modelu ‘’uruchomienia czteronożnego’’ skok inicjalizacyjny jest zdominowany przez siłę kończyn przednich i mięśni piersiowych (stanowiących ~20% – 25% całkowitej masy ciała), które wytwarzały stosunkowo dużą prędkość preludialną. Chociaż toczy się dyskusja na temat tego, czy gigantyczne pterozaury mogły wzbijać się w powietrze, powszechnie przyjmuje się, że gdyby posiadały taką umiejętność ich głównym sposobem podróżowania byłoby raczej szybowanie termiczne (które polega na wykorzystaniu prądów wstępujących do wznoszenia się a następnie szybowania) niż zrównoważonego lotu trzepoczącego, który wymagałby od Hatzegopteryx thambema ingerenci ruchu beztlenowego i byłby problematyczny do utrzymania przez dłuższy czas (obligowało by to zwierzę do utraty zbyt dużych zasobów energetycznych do wygenerowania wystarczającej siły nośnej związanej z obciążeniem skrzydeł względem regresji masy ciała, aby nie tracić wysokości i lokalizowania silnych prądów termicznych). Można więc reprezentować pogląd, iż drapieżnik tej wielkości, który wymagał względnie dużych pokładów prowiantowych mógł przemieszczać się z wyspy na wyspę, natomiast mało prawdopodobne wydają się twierdzenia z badań z wcześniejszych lat, w których wskazywano zdolności Azdarchidae do lotów bez międzylądowań przekraczających ~16093 km i bezprecedensową zdolność lotów międzykontynentalnych.
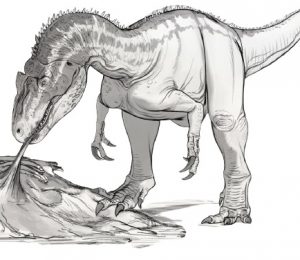
· W następstwie coraz to nowszych rekonesansów nad cechami anatomicznymi maksymalnych Azdarchidae ‘’naświetlane’’ jest to, że ich struktura szkieletowa więcej niż mniej rekomenduje przystosowanie do zerowania na lądzie do czego przychylają się także coraz to wyższe szacunki masowe niż wcześniej oczekiwano. Czynniki te uwyraźniają, że Hatzegopteryx nie unosił się wysoko nad wyspą w poszukiwaniu ofiar, a był bardziej ‘’wykfalifikowanym’’ wędrownym łowcą trzymającym kończyny bezpośrednio pod ciałem, co zapewniało mu wyprostowana postawę, a w przeciwieństwie do swoich mniejszych pobratymców jego długie kończyny gwarantowały mu dostateczną dynamikę (maksymalna wartość osiąganej prędkości to ~31km/h) za małą i średnio formatową fauną występującą na wyspie Hateg (jak choćby ornitopod Zalmoxes robustus czy nodozauryd Struthiosaurus transylvanicus). Ta adaptacja wspólnie z ewolucyjnym wyspecjalizowaniem anatomicznym w postaci stosunkowo krótkiej (wzór regresji wskazuje, że zestaw kręgów szyjnych holotypu Hatzegopteryx thambema mierzyłyby ~1,61 metra. Tymczasem te same u Quetzalcoatlus northropi TMM 41450 -3 nobilitują na ~2,86 metra), masywnej, rozciągliwej, muskularnej i odpornej na wpływy mechaniczne szyi (kręgi szyjne Hatzegopteryx były w stanie wytrzymać maksymalnie 10 krotnie większe natężenie sił zrywających niż u Arambourgiania), niemalże 2,23 metrową czaszką z mocnymi przyczepami mięśniowymi, pogrubionym kośćcem czyniły z Hatzegopteryx thambema ucieleśnienie niebezpiecznego i skutecznego myśliwego operującego bardziej brutalną strategią łowiecką charakteryzującą się używaniem potężnego dzioba do uderzająco – dźgających ruchów, aby poradzić sobie z praktycznie każdym spektrum potencjalnych ofiar egzystujących w osobliwej ekosferze wyspiarskiej (szczególnie jeśli przypiszemy dla Hatzegopteryx analogię do teraźniejszych lądowych ptaków drapieżnych jak choćby Leptoptilos crumenifer – Marabut afrykański) oraz znacznie bardziej zróżnicowanego pod względem kształtu i ekologii niż wcześniej sądzono.

Współczesna idea gigantycznego pterozaura z rodziny Azdarchidae którego przedstawicielem jest Hatzegopteryx thambema jako dominującego drapieżnika lądowego, zdolnego do polowania i konsumpcji małych i średnich zwierząt w ekosferze jego bytowania. Tutaj osobnik na pierwszym palnie z monumentalnymi proporcjami łba, dzioba i ciała (jak na wzorce pterozaurów) wykorzystując element zaskoczenia usiłuje upolować młode osobniki gatunku Telmatosaurus transylvanicus. Ta ekologiczna scena jest tylko hipotetyczna, ale z pewnością nie musiała być odległa od 71 – 68 milionowej rzeczywistości. Kreator statuetki: Mu Xi, wersja ubarwienia: Tison Zhang.
Bibliografia użyta przy realizacji artykułu:
· Grigorescu, D (1983). A stratigraphic, taphonomic and paleoecologic approach to a “forgotten land”: the dinosaur-bearing deposits of the Hateg Basin (Transylvania-Romania). Acta Palaeontologica Polonica 28, 103-121.
· Buffetaut, E., Grigorescu, D., & Csiki, Z. (2002). A new giant pterosaur with a robust skull from the latest Cretaceous of Romania. Naturwissenschaften, 89(4), 180-184, doi:10.1007/s00114-002-0307-1
· Buffetaut, E.; Grigorescu, D.; Csiki, Z. (2003). „Giant azhdarchid pterosaurs from the terminal Cretaceous of Transylvania (western Romania)” Geological Society, London, Special Publications. 217 (1): 91–104, doi:10.1144/GSL.SP.2003.217.01.09
· Sankar Chatterjee, R. J. Templin (2004). ‘’ Posture, locomotion, and paleoecology of pterosaurs’’. ResearchGate.net DOI:10.1130/0-8137-2376-0.1
· Grigorescu, D. (2005). „Rediscovery of a ‚forgotten land‘: The last three decades of research on the dinosaur-bearing deposits from the Hațeg Basin“. Acta Palaeontologica Romaniae 5: 191–204.
· Bojar, A.V., Grigorescu, D., Ottner, F., and Csiki, Z. (2005). Palaeoenvironmental interpretation of dinosaur– and mammal-bearing continental Maastrichtian deposits, Haţeg basin, Romania. Geological Quarterly 49, 205-222.
· Therrien, F. (2006). Depositional environments and fluvial system changes in the dinosaur-bearing Sânpetru Formation (Late Cretaceous, Romania): post-orogenic sedimentation in an active extensional basin. Sedimentary Geology 192, 183-205.
· Csiki, Z., Ionescu, A., and Grigorescu, D. (2008). The Budurone microvertebrate fossil site from the Maastrichtian of the Haţeg Basin – flora, fauna, taphonomy and paleoenvironment. Acta Palaeontologica Romaniae 6, 49-66.
· Michael B. Habib (2008). ‘’ Comparative evidence for quadrupedal launch in pterosaurs’’.
· Mark P. Witton ,Darren Naish (2008). ‘’ A Reappraisal of Azhdarchid Pterosaur Functional Morphology and Paleoecology’’. PolosOne, https://doi.org/10.1371/journal.pone.0002271
· Leon P.A.M Claessens, Patrick M.O’Connor,David M.Unvin (2009). „Respiratory Evolution Facilitated the Origin of Pterosaur Flight and Aerial Gigantism „.PolosOne. https://doi.org/10.1371/journal.pone.0004497
· Katsufumi Sato, Kentaro Q. Sakamoto, Yutaka Watanuki, Akinori Takahashi, Nobuhiro Katsumata, Charles-André Bost, Henri Weimerskirch (2009). „Scaling of Soaring Seabirds and Implications for Flight Abilities of Giant Pterosaurs „. PolosOne https://doi.org/10.1371/journal.pone.0005400
· Benton, M.J., Csiki, Z., Grigorescu, D., Redelstorff, R., Sander, P.M., Stein, K., and Weishampel, D.B. (2010). „Dinosaurs and the island rule: The dwarfed dinosaurs from Hațeg Island. Archived 2011-07-10 at the Wayback Machine” Palaeogeography, Palaeoclimatology, Palaeoecology, 293(3-4): 438–454
· Csikia, Z.; M. J. Bentonb (2010). „An island of dwarfs — Reconstructing the Late Cretaceous Hațeg palaeoecosystem„. Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (3–4): 265–270. doi: 10.1016/j.palaeo.2010.05.032
· Stein, K.; Csiki, Z.; Rogers, K. C.; Weishampel, D. B.; Redelstorff, R.; Carballido, J. L.; Sander, P. M. (2010). „Small body size and extreme cortical bone remodeling indicate phyletic dwarfism in Magyarosaurus dacus (Sauropoda: Titanosauria)”. Proceedings of the National Academy of Sciences. 107 (20): 9258–9263
· Codrea A. Vlad, Cătălin Jipa, Zoltan Csiki-Sava, Cristina Fărcaş, Vremir Matyas, Godefroit Pascal, Thierry Smith (2010). „More than just Nopcsa’s Transylvanian dinosaurs: A look outside the Haţeg Basin„. Palaeogeography, Palaeoclimatology, Palaeoecology. 293 (3–4): 391–405. doi: 10.1016/j.palaeo.2009.10.027
· Mark P.Witton, Michael B.Habib (2010). ‘’On the Size and Flight Diversity of Giant Pterosaurs, the Use of Birds as Pterosaur Analogues and Comments on Pterosaur Flightlessness’’. Polos One. Doi.org/10.1371/journal.pone.0013982
· Donald M. Henderson (2010). ‘’ Pterosaur Body Mass Estimates from Three-Dimensional Mathematical Slicing’’. Journal of Verterbrate Paleontology . DOI:10.1080/02724631003758334
· David B Weishampel, Coralia-Maria Jianu (2011) ‘’ Transylvanian Dinosaurs’’
· Weishampel, David B.; Butler, Richard; Prondvai, Edina; Ősi, Attila (2012). „Phylogeny, Histology and Inferred Body Size Evolution in a New Rhabdodontid Dinosaur from the Late Cretaceous of Hungary„. PLOS ONE. 7 (9): e44318. doi: 10.1371/journal.pone.0044318
· Dave Weishampel, Oliver Kerscher (2013). „Franz Baron Nopcsa: A short life of research in dinosaur evolutionary paleobiology and Albanology „. ResearchGate. DOI:10.1080/08912963.2012.689745
· Mark P. Witton and Darren Naish. (2015). ” Azhdarchid pterosaurs: water-trawling pelican mimics or terrestrial stalkers „. Acta Palaeontologica Polonica. doi:http://dx.doi.org/10.4202/app.00005.2013
· Zoltan Csiki-Sava, Eric Buffetaut, Attila Ősi, Xabier Pereda Suberbiola (2015). ‘’ Island life in the Cretaceous – Fauna Island life in the Cretaceous – Faunal composition, biogeography, evolution, and extinction of landliving vertebrates on the Late Cretaceous European archipelago’’. National Center for Biotechnology Information. doi: 10.3897/zookeys.469.8439
· Mark P.Witton (2016). ‘’The lives and times of flying reptiles as told by the fossil record, part 1: azhdarchid pterosaurs’’. Markwitton.com Blog
· Naish, D.; Witton, M.P. (2017). „Neck biomechanics indicate that giant Transylvanian azhdarchid pterosaurs were short-necked arch predators„. PeerJ. 5: e2908. doi:10.7717/peerj.2908
· Witton, M.P. (2017). „New paper: when the short-necked, giant azhdarchid pterosaur Hatzegopteryx ruled Late Cretaceous Romania”. Markwitton.com Blog
· Mátyás Vremir, Gareth Dyke, Zoltán Csiki-Sava, Dan Grigorescu, Eric Buffetaut (2018). ” Partial mandible of a giant pterosaur from the uppermost Cretaceous (Maastrichtian) of the Hațeg Basin, Romania „. Researgate.net, DOI:10.1111/let.12268
· Larramendi, A.; Paul, G. S.; Hsu, S.-y. (2020). A Review and Reappraisal of the Specific Gravities of Present and Past Multicellular Organisms, with an Emphasis on Tetrapods. The Anatomical Record. doi: https://doi.org/10.1002/ar.24574
· Ana Benítez-López, Luca Santini, Juan Gallego-Zamorano, Borja Milá, Patrick Walkden, Mark A. J. Huijbregts & Joseph A. Tobias (2021). ”The island rule explains consistent patterns of body size evolution in terrestrial vertebrates” / Nature Ecology & Evolution. 2021. DOI: 10.1038/s41559-021-01426-y.
· Goto, Yusuke; Yoda, Ken; Weimerskirch, Henri; Sato, Katsufumi (2022). „How did extinct giant birds and pterosaurs fly? A comprehensive modeling approach to evaluate soaring performance”. PNAS Nexus. 1 (1): pgac023. doi:10.1093/pnasnexus/pgac023.
· Peter Menszikov (2023). ‘’ Flying abilities and mass calculation of giant Azhdarchids’’. Milset ESI – International Movement for the Promotion of Science and Technology.
Treść merytoryczna opublikowanego artykułu na łamach witryny Dinoanimals.pl może być wykorzystywana jedynie na własny użytek edukacyjny i nie może stanowić podmiotu informacyjnego dla innych blogów. Wykluczonym jest także udostępnianie ani publikowanie go poprzez kanały tematyczne bez zgody jego autora.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

Cesarz, król i królowa
Poprzedni artykuł:
Cesarz, król i królowa Następny artykuł:
”Intelektualiści” ?
Cesarz, król i królowa Następny artykuł:
”Intelektualiści” ?





































Ciekawe jest, że niektóre dinozaury nie mają nazw w języku polskim (chociażby wyżej wymieniony Hatzegopteryx), a mogłyby. Spotkałem się kiedyś z określeniem „Drednotus”, odnośnie Dreadnoughtusa (taki duży zauropod nazwany na cześć pancernika Drednot. Oczywiście nikt nie uznaje tego za oficjalną nazwę polską. Z kolei gdyby spróbować spolszczyć ⬆️TO PTASZYSKO, to wyszedłby nam Hacegopteryks…