Dilophosaurus został po raz pierwszy odkryty przez w 1940 roku przez Jessa Williamsa z plemienia Navajo, materiał skalny tego teropoda wydobyty został natomiast dwa lata później przez zespól paleontologów Sama Wellsa i Wyana Longstona Jr z University of California Museum of Paleontology, którzy tym samym przyczynili się do ‘’ujrzenia światła dziennego’’ przez tego największego dinozaura drapieżnego jaki został dotąd odkryty w Ameryce Północnej. Jego wczesne odkrycie miało ogromne znaczenie dla naszego zrozumienia ewolucji pierwszych dużych teropodów, mimo to Dilofozaur nigdy nie został dokładnie opisany naukowo do tej pory. Przez ostatnie sześć lat kooperacja dwóch naukowców Adama D.Marsha oraz Timothego B. Rowa bardzo ciężko pracowała nad opisywaniem i analizowaniem każdej kości każdego znanego szkieletu przypisywanemu dilofozaurowi w tym kilka nowych okazów.
Nowa analiza autorów pozwoliła określić o wile bardziej precyzyjnie, gdzie w drzewie genealogicznym znajduje się Dilofozaur, pierwotnie uznano go za gatunek teropoda z rodziny Megalozaurów, było tak aż do momentu odkrycia słynnego wysokiego podwójnego grzebienia kostnego, wówczas zmieniono jego nazwę i zinterpretowano go jako swoistego rodzaju pośrednika między małymi celofizoalnymi teropodami triasowaymi a późniejszymi jurajskimi ceratozaurami. Lecz analiza Adama i Timothego ujawnia coś fascynującego mianowicie to, iż Dilofozaur nie jest spokrewniony z żadnym z tych zwierząt zamiast tego tworzy dziwną klasę teropodów wraz z Zupaysaurus rougieri z Ameryki Południowej i Crylophosaurus ellioti z Antarktydy i żaden z tych dinozaurów nie był bezpośrednim przodkiem ceratozaurów lub megalozaurów z późnej Jury.

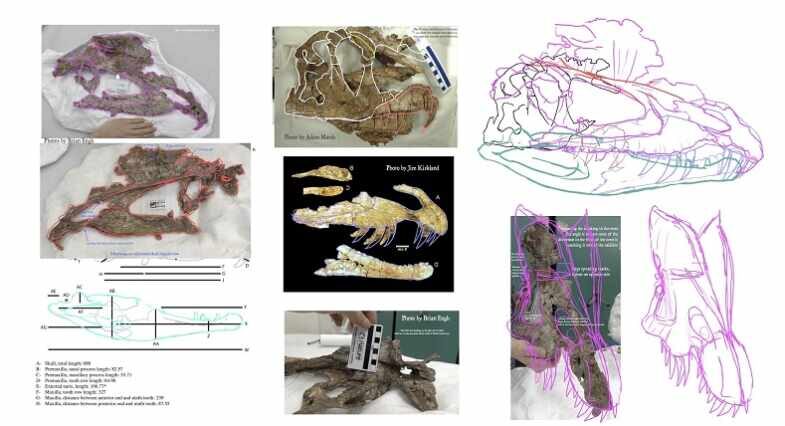
Fotografia ukazująca badany przez Adama D.Marsha oraz Timothego materiał skalny Dilophosaurus wetherilli.
Chociaż może wydawać się to zaskakujące lub mylące należy pamiętać o tym, że od czasów Dilofozaura teropody były już dinozaurami odnoszącymi duże sukcesy globalne od milionów lat, po prostu na chwile obecną nie mamy wszystkich szkieletów obrazujących tego jak wyglądały, także analiza Adama i Thimothego potwierdzają ewolucje dużych rozmiarów ciała u dinozaurów nie był uporządkowaną liniową progresją od małych dinozaurów z okres triasu do słynnych późno jurajskich gigantów, ale raczej okres wczesno jurajski był czasem ‘’dzikiego’’ zróżnicowanie ewolucyjnego i eksperymentalnego po zakończeniu triasu.
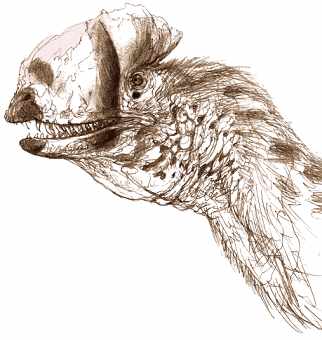
Fragmentaryczność znanych czaszek Dilofozaurów doprowadziła w przeszłości Sama Wellsa do spekulacji, iż drapieżnik ten ma delikatną anatomicznie czaszkę, z kolei ta sugestia była następstwem sugerowanego słabego nacisku szczęk co doprowadziło do dalszej spekulacji, że musiał być on padlinożercą, specjalizować się w diecie opartej na rybach lub ‘’ujarzmiać’’ swoją potencjalną ofiarę jadem jak zostało to przedstawione w ‘’Parku Jurajskim’’. Aby lepiej zrozumieć budowę anatomiczną i biologię Dilofozaura wyżej wymienieni autorzy podjęli współpracę z paleoartystą i paleontologiem amatorem przy tworzeniu najbardziej aktualnej i rygorystycznej formie odwzorowania czaszki tego drapieżnika, gdzie odwzorowanie obejmowało wszystkie znane materiały kopalne czaszek, biorąc pod uwagę ich trójwymiarowe struktury.
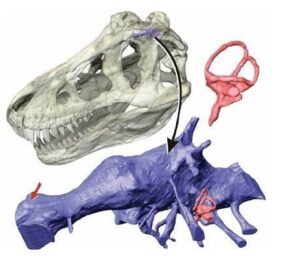
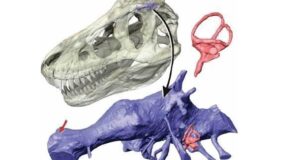
Powyższa rekonstrukcja czaszki Dilophosaurus wetherilli została stworzona przez dokładne porównania, fotografie i zestawienia wszystkich znanych okazów w celu uzyskania najbardziej rygorystycznej pełnej rekonstrukcji czaszki tego teropoda. Brakujące części grzebienia kostnego sugeruje tylko częściowa sylwetka, prawdziwy rozmiar jak i kształt grzebienia są spekulatywne.
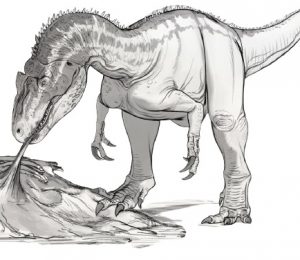
Ta dogłębna analiza wszystkich znanych materiałów skalnych czaszek Dilofozaura szybko wyjaśniła, że wszystkie wcześniejsze hipotezy na temat biologii Dilofozaura są całkowicie bezpodstawne. Najdokładniejsza jak dotąd rekonstrukcja i skalowanie ujawniły, że szczęki Dilofozaura były rzeczywiście głębokie i silnie umięśnione (autorzy wspominają o naprawdę imponującym obszarze przyczepu mięśni), a jego zęby były duże i ząbkowane idealnie przystosowane do przecinania mięsa i kości. Dilofozaur jak się okazuje był więc aktywnie polującym zabójcą innych zwierząt dzielących z nim ekosystem i największym drapieżnikiem w swoim środowisku, teorię te wspierają ślady ugryzień na kościach Sarahsaurus aurifontalis (TMM 43646 – 2.56).

Szkic przedstawiający sekwencje ataku Dilophosaurus wetherilli na Sarahsaurus aurifontalis autorstwa paleoartysty Briana Engh’a.
Kilka badanych okazów Dilofozaurów ma liczne urazy i ślady ugryzień, które wykazują oznaki wyleczenia, co sugeruje na brutalny styl życia i agresywność, która prowadziła do walk wewnątrzgatunkowych a nawet kanibalizmu tak samo jak ma to miejsce u wielu jego współczesnych krewnych ptaków.

Agresja wewnątrzgatunkowa była domena wielu teropodów, ślady ugryzień na badanym materiale skalnym Dilophosaurus wetherilli sugerują, że brutalne ataki i walki nie były odosobnione w przypadku tego teropoda zarówno podczas walk terytorialnych, godowych czy o pożywienie. Nie jest wykluczonym, iż w przypadku Dilophosaurus panował kanibalizm w celu eliminacji późniejszych oponentów – szkic autorstwa Briana Engh’a.
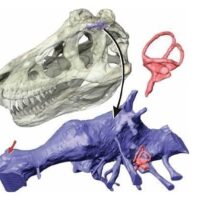
Dilofozaur ma jednej z najbardziej dziwacznych i wyróżniających się grzebieni kostnych pośród dinozaurów, który do tej pory był niezbyt rozumiana cechą anatomiczna i w większości przedstawień w paleoartach przedstawiany był jako pokryty cienką warstwą skórną i oddzielony anatomicznie przez okno przedoczodołowe, autorzy publikacji jednak badając szczegółowo czaszki dają nam jasno do zrozumienia, iż herb tego teropoda nie jest w ogóle oddzielony od okna przedoczołodowego a jest jej rozwinięciem. W rzeczywistości większość teropodów ma małe grzebienie utworzone przez kość łzową i kość nosową w tym samym obszarze i prawie wszystkie z tych grzebieni wewnątrz mają puste przestrzenie pozostawione przez kieszenie powietrzne (nie mylić z workami powietrznymi) wychodzące okna przedoczodołowego aż do grzebienia kostnego. Grzebień Dilofozaura doprowadza ten wzorzec rozwojowy do dziwacznej skrajności rozszerzając przestrzenie powietrzne tak bardzo, że nawet nie jest możliwym ocenienie pełnego rozmiaru i kształtu grzebienia kostnego tego drapieżnika, ponieważ górna krawędź nie jest zachowana. Z powodu bocznego zgnioty skamieniałości nie jest również jasne, ile warstwy cienkościennej kości wypełnionej powietrzem faktycznie tworzy każdą z nich, ‘’żywa kość’’ jest o wiele trwalsza niż krucha skamieniałość, poza tym w materiale skalnym obejmującym grzebień brakuje większego obszaru lub jest on zmiażdżony, więc trudnym jest do oszacowania jak mocna byłaby sama kość.

Szkic ukazujący wewnętrzną budowę anatomiczną grzebienia kostnego Dilophosaurus wetherilli ukazująca fakt iż herb tego drapieżnika nie był oddzielony od okna przedoczodołowego a w rzeczywistości był jego rozwinięciem – autor Brian Engh.
Grzebień Dilofozaura był znacznie trwalszy z innego powodu , wierzcie lub nie tylko wymarłe dinozaury mają wypełnione kieszeniami powietrznymi kościste grzebienie. W rzeczywistości istnieją dziesiątki gatunków dinozaurów o podobnych herbach, które wciąż żyją. Wiele współczesnych ptaków ma również wypełnione kieszeniami powietrznymi kostne grzebienie połączone z zatokami, a praktycznie u wszystkich grzebienie okryte są keratyną, tym samym twardym materiałem, z którego składają się ptasie dzioby. Keratyna jest niezwykle wytrzymałym materiałem do tego stopnia, iż dorosłe współczesne Kazuary mogą bezpiecznie biegnąć z maksymalną dla nich prędkością przez gęstą dżunglę, a niektóre z samców dużych gatunków Dzioborożców (Bycanistes bucinator, Buceros bicornis, Buceros rhinoceros) walczą ze sobą w okresie godowym przy użyciu grzebieni. Niektóre z tych herbów Dzioborożców ledwo mają podtrzymujące je kości (Buceros hornbill), w ogóle keratynowe pokrycie również znacznie powiększa grzebień u niektórych ptaków, keratyna rozszerza rozmiar herbu do 1,5 razy większego rozmiaru (Numida meleagris). Keratyna z grzebieni kostnych (Aceros cassidix, Crax rubra, Rhabdotorrhinus corrugatus) czy dziobów może przybierać również dowolne kolory (Ramphastos sulfuratus, Ramphastos toco), dlatego to efektowne dzioby i grzebienie odgrywają ważną rolę w rozpoznawaniu gatunków i pokazach seksualnych. Pomagają ptakom również w komunikacji i przyciąganiu partnerów, ale pełnią też inne funkcje, jedno z ostatnich badań wykazało, że grzebień Kazuara jest ważną strukturą do regulacji termicznej. Gdzie mogą one za jego pośrednictwem odprowadzać nadmiar ciepła co pomaga utrzymać ich mózg w odpowiedniej temperaturze w gorącym tropikalnym klimacie, w którym żyją.
Jedną z najciekawszych hipotez w pracy autorów jest to, iż Dilofozaur mógł się cechować rodzajem nadmuchiwanego worka skórnego podobnego do dzisiejszych Bucorvus leadbeateri, Fregata magnificens, Phalacrocorax carbo czy Pelecanus onocrotalus, które są połączone u tych gatunków z płucami co pozwala na bardziej skuteczną absorpcję tlenu przy jednoczesnym zatrzymaniu wody podczas oddychania czy wzmocnieniu wokalizacji.

Model pacynki Dilophosaurus wetherilli wykonany przez paleoartystę Briana Engh’a ukazujący nadmuchiwane worki skórne teropoda.
Wiele z kości Dilofozaura oraz ozdoby na łbie drapieżnika było zaopatrzonych w wspomniane przeze mnie w artykule kieszenie powietrzne ta adaptacja prawdopodobnie pomogła w umożliwieniu Dilofozaurowi i późniejszym teropodom uzyskanie dużych rozmiarów, a jednocześnie skomplikowane przestrzenie powietrzne w ich kościach pomogły mniejszym teropodom stać się wystarczająco lekkimi by mieć możliwość wzbijania się w powietrze. Ekspansja kieszeni powietrznych w grzebieniach Dilofozaura były jednym z głównych czynników napędzających ewolucję, osiągnięcie dużych rozmiarów u Dilofozaura byłoby więc wtórną adaptacją kieszeni powietrznych w kościach tego dinozaura.
Wyszukane funkcje ekspozycyjne takie jak efektowny keratynowy grzebień często ewoluuje u grup zwierząt, które są szeroko rozpowszechnione i odnoszą duże sukcesy, a Dilofozaur z pewnością odniósł sukces, jego szkielety zostały znalezione w wielu poziomach formacji Kayenta co sugeruje, że zwierzęta te przetrwały w Arizonie Północnej około 5,4 mln lat.
Niestety fragmentaryczny charakter zachowanych kilku czaszek Dilofozaurów sprawia, że niemożliwym jest zobaczenie jak ewoluowały ich grzebienie i jaki miały stopień zmienności, lecz opierając się na badaniach przeprowadzanych na współczesnych ptakach z podobnymi tworami można zasugerować, iż ewoluowały stosunkowo szybko i wykazywały duży stopień zmienności między blisko spokrewnionymi gatunkami a nawet wśród osobnikami tego samego gatunku. Obecność i zmienność grzebieni i kieszeni powietrznych w grupach wczesnych teropodów jurajskich takich jak omawiany Dilofozaur przyczynia się do powstania obrazu wczesnej Jury jako epoki radykalnej ewolucji łączącej w sobie eksperymentowanie i dywersyfikację a w następstwie koniec triasowego wymierania, które oznacza, że wciąż musimy się wiele nauczyć na temat tajemniczego okresu jakim była wczesna Jura jaki i ekologii, i dinozaurach z tamtego okresu, które dostosowując się do zmieniającego się klimatu przygotowały grunt pod pojawienie się największych zwierząt lądowych zarówno roślinożernych jak i mięsożernych jakie kiedykolwiek żyły na naszej planecie.
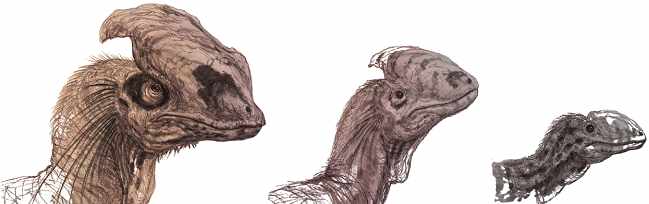
Szkic przedstawiający ontogenezę gatunku Dilophosaurus wetherilli oraz hipotetyczny rozwój kształtu i wielkości grzebienia kostnego tego teropoda – autor Brian Engh.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Poprzedni artykuł:
Prognathodon Następny artykuł:
Największy znany (obecnie) ślad teropoda
Prognathodon Następny artykuł:
Największy znany (obecnie) ślad teropoda