Mosasauridae to rodzina dużych morskich „gadów”, obecnie zawierająca 39 rodzajów stanowiących bardzo wyspecjalizowaną grupę jaszczurek (Lacertilia), które przeszły na morski styl życia. W rodzinie wyróżnia się kilka podrodzin z reguły blisko spokrewnionych – Mosasaurinae i Halisaurinae, a także Russellosaurina (w tym takie grupy, które wcześniej uważano za izolowane takie jak: Tethysaurinae, Yaguarasaurinae, Tylosaurinae i Plioplatecarpinae – Simões i współautorzy, 2017). Pierwsze szczątki kopalne przedstawiciela Mosasauridae– Mosasaurus hoffmani zostały oficjalnie odkryte w kamieniołomie wapienia w Maastricht (miasta położonego w południowej Holandii) nad rzeką Mozą w 1764 roku, co dało nazwę tej grupie (Mulder,2004). W ciągu ostatnich 20 milionów lat kredy (okres geologiczny turono – mastrychtu) wraz ze zniknięciem Ichthyosauria i upadkiem Plesiosauria, Mosasauridae stały się główną grupą dużych drapieżników morskich o pokaźnej różnorodności ekologicznej. Wyginęły w wyniku wymierania kredowego około 66 milionów lat temu (Everhart,2005), – * chociaż istnieją doniesienia o odkryciach z okresu paleogenu (Gallagher i współautorzy,2012).

Model przedstawiający rekonstrukcję wizualno-kolorystyczną Mosasaurus hoffmani pierwszego odkrytego i zarazem największego przedstawiciela Mosasauridae – autor Goro Furtura.
Ewolucja:
Mosasauridae prawdopodobnie wyewoluowały z wymarłej grupy wodnych jaszczurek wczesnokredowych znanych jako Aigialosauridae. Szkielet wczesnego Mosasauridae – Dallasaurus tureneri z podrodziny Mosasaurinae (opisany przez Bella i Polcyna, 2005) ma wiele cech obecnych w szkieletach zaawansowanych przedstawicieli podrodziny, a także w szkieletach rzekomych przodków Mosasauridae takich jak Aigialosaurus dalmaticus / Aigialosaurus bucchichi. Dallasaurus zachowywał kończyny nadające się do opcjonalnego poruszania się na lądzie, podobne w strukturze do kończyn aigialozaurów i jaszczurek lądowych (stan plezjopedyczny kończyn) w przeciwieństwie do zaawansowanych Mosasauridae, które przekształciły kończyny w płetwy (stan hydropedalny kończyn). Jednak Dallasaurus miał kilka cech naraz, które łączyły go z pochodnymi członkami podrodziny Mosasaurinae. Analiza filogenetyczna przeprowadzona przez Bella i Polcyna,2005 wykazała, iż hydropedalne Mosasauridae nie tworzyły kładu, który obejmował by taksonów plezjopedycznych takich jak: Dallasaurus, Yaguarasaurus, Russellosaurus, Tethysaurus, Haasiasaurus i Komensaurus. Analiza wykazała, że stan hydropedalny kończyn rozwijał się niezależnie w trzech różnych grupach Mosasauridae – Halisaurinae, Mosasaurinae i grupie zawierającej podrodziny Tylosaurinae i Plioplatecarpinae, wyniki tego badania filogenetycznego zostały w dużej mierze potwierdzone przez analizy przeprowadzone przez Caldwella i Palca, 2007 oraz Leblanca i współautorów,2012. Dalsze analizy filogenetyczne prowadzone przez Makádi i współautorów,2012 wskazały, że kończyny hydropedalne ewoluowały w dwóch grupach Mosasauridae (w Mosasaurinae i kładzie zawierającym Halisaurinae , Tylosaurinae i Plioplatecarpinae). Jeśli wziąć by hipotezę Bella i Polcyna,2005 za prawidłową to rodzina Mosasauridae w tradycyjnym sensie jest w rzeczywistości grupą polifiletyczną, w swojej analizie wcześniej wymienieni autorzy poparli również monofilię Mosasauridae poprzez włączenie do rodziny Dallasaurus i innych wyżej wymienionych taksonów plezjopedycznych, podczas gdy Caldwell,2012 nieoficjalnie zaproponował ograniczenie rodziny Mosasauridae tylko do samego Mosasaurus i jego najbliższych hydropedalnych krewnych.

Model przedstawiający rekonstrukcję wizualno – kolorystyczną Dallasaurus tureneri. Dallasaurus stanowił fascynujące odkrycie, gdyż postrzegany jest jako forma przejściowa, ponieważ jego ciało i ogon wykazują adaptację do pływania, podczas gdy kończyny miały wciąż większy udział w rozwoju przystosowawczym do poruszania się na ladzie.
Dokładna pozycja filogenetyczna kładu zawierającego rodzinę Mosasauridae i ich bezpośrednich krewnych ( Aigialosauridae i Dolichosauridae ) w obrębie łuskonośnych (Squamata) pozostaje niejasna. Niektóre analizy kładystyczne stwierdzają, że są oni najbliższymi krewnymi węży biorąc pod uwagę podobieństwa w anatomii dolnej szczęki i czaszki (Lee,1997), jednak zostało to zakwestionowane, a analiza morfologiczna przeprowadzona przez Conrada,2008 umieściła Mosasauridae jako przedstawicieli nadrodziny Varanoidea – blisko spokrewnionych z współczesnymi wronowatymi (Varanidae). Późniejsza eksplikacja związków filogenetycznych w obrębie Anguimorpha przeprowadzona przez Conrada i współpracowników, 2011 opierająca się wyłącznie na morfologii umieściła Mosasauridae , Aigialosauridae i Dolichosauridae jako Anguimorpha, leżąc po za kładem zawierającym waranowate (Varanidae) i helodermowate (Helodermatidae). Z drugiej strony zestawienie oparte na połączonych zestawach danych badań morfologicznych i molekularnych wykazały, że Mosasauridae są bliżej spokrewnione z Varanidae i Lanthanotidae niż z Helodermatidae oraz Shinisauroidea (Conrad i współautorzy,2011). Obszerna analizamorfologiczna Gauthiera i współpracowników,2012 umieszcza Mosasauridae , Aigialosauridae i Dolichosauridae w nieoczekiwanej pozycji jako podstawowych przedstawicieli Scincogekkonomorpha (kłady zawierające wszystkie taksony, które są morfologicznie bliższe gekonowatym i scynkowatym niż legwanowatym), które nie zostały uwzględnione w kładzie Scossrog. Pozycja filogenetyczna tych taksonów okazała się bardzo zależna od tego, które taksony zostały uwzględnione lub wyłączone z analizy. Gdy Mosasauridae zostały wyłączone z analizy Aigialosauridae i Dolichosauridae wkroczyły do Scleroglossa tworząc siostrzana grupę (w tym Serpentes, Amphisbaenia, Dibamidae i Anniellidae). Natomiast gdy w eksplikacji uwzględniono Mosasauridae, a różne taksony ze zmniejszonymi lub brakującymi kończynami zostały wykluczone z wyjątkiem węży (na przykład Amphisbaenia i Dibamidae) – Mosasauridae , Aigialosauridae i Dolichosauridae weszły do Scleroglossa tworząc siostrzaną grupę z wężami. Longrich i współpracownicy,2012 przeprowadzili analizę morfologiczną filogenetycznych związków łuskowatych, wykorzystując dane Gauthiera i współautorów,2012 stwierdził, że filogenetyczna pozycja kładu zawierającego Mosasauridae i ich bezpośrednich krewnych w Squamata jest bardzo niestabilna, a grupa jest poza Scleroglossa lub obok form, które utraciły kończyny. Z drugiej strony współczesne badania pokazują, że taksony Scleroglossa i Scincogekkonomorpha nie są ważne, ponieważ tradycyjnie oddzielone łuskowate legwany w rzeczywistości są spokrewnione z wężami i Anguimorpha tworząc z nimi kład Toxicofera (Pyron i współpracownicy,2013). Tak więc pozycja rodziny Mosasauridae i pokrewnych grup Squamata w tej chwili najbardziej niezawodnie odzwierciedla połączoną analizę Conrada i współpracowników.

Kładogram Mosasauridae autorstwa Simõesa i współautorów, 2017.
Ogólna charakterystyka:
Badanie izotopów tlenu w zębach Mosasauridae autorstwa Bernarda i współautorów,2010 wykazało, że rozwinięci członkowie rodziny mieli zwiększone tempo metabolizmu w porównaniu do typowych ektoterm i miały standardową temperaturę ciała w okolicach 35 ± 2 – 39 ± 2 °С. Jednak w tym badaniu nie wykorzystano powiązanych skamielin zwierząt ciepłokrwistych do porównania izotopów tlenu, ale dane nie wykluczają, że temperatura ciała Mosasauridae może nadal być częściowo zależna od temperatury otoczenia. Mikrostruktura tkanki kostnej Mosasauridae odpowiada pośredniemu poziomowi metabolizmu pomiędzy występującemu współcześnie Dermochelys coriacea (żółw skórzasty) a Ichthyosauria lub Plesiosauria, których ciepłokrwistość i bardzo wysokie tempo metabolizmu są obecnie uważane za udowodnione (Houssaye, 2013). Tak więc Mosasauridae nie mogły być całkowicie ciepłokrwiste, ale po prostu miały rozwinięto formę mezotermii. Dalsze analizy izotopowe przeprowadzone przez Harella i współpracowników,2016 wykazały, że Mosasauridae były ciepłokrwiste, porównując skład izotopowy trzech kampańskich rodzajów (Clidastes, Tylosaurus i Platecarpus) z danymi znanymi dla zimnokrwistych ryb z rodzaju Enchodus i z pewnością ciepłokrwistych ptaków z rodzaju Ichthyornis, autorzy doszli do wniosku, iż Mosasauridae mają wewnętrzny mechanizm termoregulacji i pozostają cieplejsze niż otaczająca je woda. Zasadniczo obliczona temperatura ciała badanych Mosasauridae była porównywalna z temperaturą ciała współczesnych ptaków morskich i była wyższa u mniejszego Platecarpus (36,3 °C) niż u większego Tylosaurus (34,3 °C), co nie pozwala na współczynnik dużej temperatury. Mosasauridae są znane z obszarów daleko wysuniętych na północnych szerokościach geograficznych (Nicholls i współpracownicy 2002; Konishi i Sues, 2012; Grigoryev, 2017) oraz z obszarów daleko wysuniętych na południowych szerokościach geograficznych (Martin,2007; Otero i współautorzy,2016), co również wskazuje na obecność rozwiniętych mechanizmów termoregulacji.
Tradycyjnie Mosasauridae były uważane za drapieżniki zasadzkowe, które charakteryzują się pływaniem typu Anguilliformes (węgorzokształtnych) to znaczy ruchem spowodowanym zaginaniem ciała, skutecznym do szybkiego przyspieszania, ale nie w stosunku do dłuższego pływania (Massare,1987). Jednak morfologia końcowych kręgów ogonowych (Lindgren, 2011) i odciski tkanek miękkich (Prognathodon sp. ERMNH HFV 197; Lindgren i współpracownicy,2013; Platecarpus tympaniticus LACM 128319; Lindgren i współautorzy, 2010) pokazują, że nawet stosunkowo wczesne Mosasauridae miały płetwy ogonowe podobne do Metriorhynchidae, Ichthyosauria i niektórych rekinów. Badanie ruchliwości poszczególnych odcinków kręgosłupa wspiera również hipotezę iż Mosasauridae były aktywnymi pływakami pelagicznymi i stosowały głównie typ poruszania się określany mianem – Carangiform (czyli taką w której mięśnie poruszające ciałem przesuwają się na trzecią część ciała – głównie ogon – Lindgren i współpracownicy, 2007; Carpenter, 2017). Dlatego zamiast aktywnie się ‘’wiercić’’ podczas ruchu przednia część Mosasauridae pozostawała względnie nieruchoma co zmniejszało opór wody. Zdecydowana większość siły lokomotorycznej opierała się bezpośrednio na ruchach ogona, zwierzęta te więc nie były wyspecjalizowanymi łowcami ‘’zasadzkowymi’’ i mogły aktywnie ścigać zdobycz w wodzie (Lindgren i współautorzy, 2010, 2011). Dane te są zgodne ze wskaźnikami metabolicznymi Mosasauridae sugerującymi aktywny tryb życia. Prawdopodobnie ogony Mosasauridae były podtrzymywane przez kil boczny, który jest charakterystyczny dla wielu aktywnie pływających zwierząt. Korelaty osteologiczne nie wspierają obecności płetwy grzbietowej u Mosasauridae (możliwym wyjątkiem może być Plotosaurus– Lindgren i współpracownicy,2007) stabilizacja ciała podczas pływania została prawdopodobnie osiągnięta głownie dzięki ruchowi kończyn przednich.
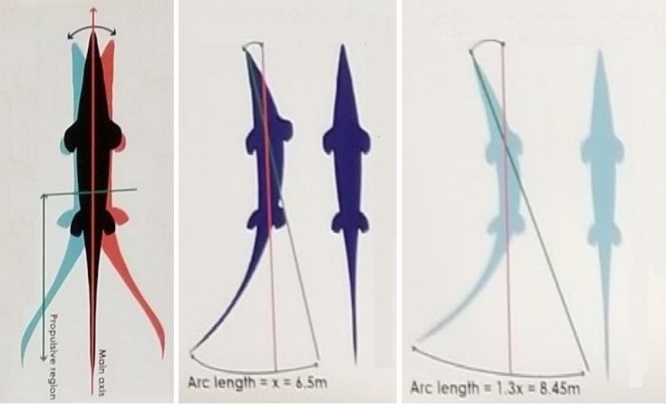
Mosasauridae prawdopodobnie używały mechanizmu lokomocji w wodzie określanego mianem Carangiform cechującym się siłą motoryczną płetwy ogonowej z lekkim bocznym ruchem łba i elastyczności napędowej pochodzącej z ostatnich 2/3 ciała o dużej amplitudzie na ogonie.
Znane są również dobrze zachowane odbitki łusek Mosasauridae, które również wykazują pewne dostosowania do aktywnego trybu życia. Materiał skalny z późnego Mastrychtu w Jordanii wykazał, że ciała Mosasauridae, a także błony między ‘’palcami’’ kończyn przednich i tylnych były pokryte małymi nakładającymi się łuskami romboidalnymi przypominającymi łuski węża. Tylosaurus proriger o sygnaturze katalogowej KUVP 1075 znaleziony w hrabstwie Gove w stanie Kansas ‘’ukazuje’’ skórę składającą się z małych łusek pod względem wielkości, kształtu i położenia bardzo przypominające te u współczesnych łuskowatych (ich gęstość osiąga około 90 płatków na cal kwadratowy). Porównanie ze skórą współczesnego Crotalus oreganus (grzechotnik pacyficzny) wykazały, że łuski były nieco mniejsze wielkościowo niż wcześniej wspomnianego węża, którego skóra zawiera średnio około 80 płatków.

Odcisk łusek skórnych Tylosaurus proriger KUVP 1075.
Podobnie jak w przypadku współczesnych jaszczurek łuski Mosasauridae mogły się nieco różnić na powierzchni ciała tych zwierząt pod względem rodzaju i wielkości. Próbka z Jordanii ukazuje co najmniej dwa rodzaje łusek: łuski kilowe pokrywające górną część ciała i łuski gładkie pokrywające dolne rejony ciała (Kuddumi,2009). Skóra Plotosaurus bennisoni (UCMP 152664) jednego z bardziej zaawansowanych przedstawicieli podrodziny Mosasaurinae zawiera łuski wzmocnione kilem i osteodermą, podobne strukturalnie łuski opisano również u bardziej prymitywnego Platecarpus tympaniticus (LACM 128319) z podrodziny Plioplatecarpinae. Taki kształt łusek rzekomo przyczyniał się do przednio – tylnego przepływu wody i zmniejszenia aktywności mikroturbulencyjnej wzdłuż powierzchni skóry (Lindgren i współpracownicy, 2009). Co ciekawe skala łusek u okazu holotypowego Ectenosaurus clidastoides jest mniejsza (2,7 x 2 mm) niż u Platecarpus tympaniticus LACM 128319 (3,8 x 4,4 mm) pomimo faktu, że zwierzęta są w przybliżeniu tej samej wielkości (Lindgren i współautorzy, 2011).

Grafika przedstawiająca strukturę powłokową jednego z najbardziej zaawansowanych Mosasauridae: Plotosaurus bennisoni (UCMP 152664) – Należy zwrócić uwagę na małe rozmiary skal przypuszczalnie dostosowanie w celu zwiększenia wydajności hydrodynamicznej. Zachowane pigmenty wskazują, że Mosasauridae były ciemne na górze i jasne na dole, jak współczesny Dermochelys coriacea – żółw skórzasty (Lindgren i współautorzy,2014).
Znane cechy wewnętrznej struktury Mosasauridae mają wiele różnic w stosunku do współczesnych jaszczurek, Platecarpus tympaniticus (LACM 128319) oprócz odcisków skóry zachowuje również odciski narządów wewnętrznych. Kilka czerwonych obszarów w skamieniałości prawdopodobnie reprezentują: serce, płuca i nerki , po za tym w wcześniej wspomnianym okazie zachowane są siatkówka oraz tchawica. Wracając do kwestii wewnętrznej budowy anatomicznej nerki ulokowane były głębiej w jamie brzusznej niż u współczesnych waranowatych – ich lokacja jest bardziej podobna do położenia waleni. Rurki oskrzelowe prowadzące do płuc u Platecarpus przechodzą równolegle do siebie jak u waleni i nie odbiegają od siebie jak u jaszczurek lądowych (Lindgren i współpracownicy,2010). Niemal na pewno Mosasauridae miały zmodyfikowane układ krążenia podobny do tych jak u współczesnych waranowatych niezbędny do zwiększenia ciśnienia krwi i zwiększonego ogólnego poziomu metabolizmu.
Kopulacja Mosasauridae mogła wyglądać jak u współczesnych delfinów – brzuch przy brzuchu, nacieranie na siebie pod różnymi kątami. Badania niektórych okazów wskazują ,że Mosasauridae rodziły żywe młode (wiadomo, iż produkcja jaj niezależnie rozwijała się jednocześnie w kilku liniach współczesnych łuskowatych) i, że przynajmniej niektórych przypadkach przypadkach młode spędzały pierwsze lata życia na otwartym oceanie, a nie w bezpiecznych płytkich obszarach wodnych (Field i współautorzy, 2015; Everhart, 2017). Materiał skalny Mosasauridae z rodzaju Plioplatecarpus zachowany został z szczątkami pięciu młodych wewnątrz jamy brzusznej, nie wykazując przy tym oznak, że zostały one zjedzone, takich jak na przykład rozpuszczenia kości przez kwas żołądkowy. Opieka nad rozwojem potomstwa u Mosasauridae wydaje się mało prawdopodobna, biorąc pod uwagę ich aktywny tryb życia i brak podobnych zachowań u współczesnych waranowatych jak i innych współcześnie żyjących łuskowatych.

Przedstawiciel rodziny Mosasauridae – Clidastes liodontus wydający na świat potomstwo – grafika autorstwa paleoartysty Juliusa Csotonyi.
Mosasauridae posiadały tekodontowaty układ dentystyczny (zęby osadzone w zębodołach szczęki i żuchwy), który nie był charakterystyczny dla współczesnych łuskowatych. Prawdopodobnie w związku z tym ich zęby nie zostawały ukryte przez mięsiste dziąsła, co obserwuje się u współczesnych waranowatych. Okres zmiany zębów u Platecarpus (JAG 0006 t) wynosił około 260 dni, u Prognathodon (JAG 0005 t) około 374 dni a u Tylosaurus (JAG 0007 t) około 593 dni. Przez cały okres swojego istnienia Mosasauridae zajmowały nisze ekologiczne średnich i dużych drapieżników, dlatego większość garnków miała morfotyp zęba tnącego w pewnym stopniu odpowiednich do ataku i zabijania potencjalnie dużej ofiary. Kolejnym najczęstszym morfotypem zęba był przeszywający odpowiedni do przytrzymywania stosunkowo małej ale ruchliwej zwierzyny łownej (Poynter, 2011; Ross, 2006).
Wcześni przedstawiciele Mosasauridae, podobnie jak ich ziemscy przodkowie miały kinetyczne czaszki, które pozwalały im na całkowite połykanie stosunkowo dużej zdobyczy (Everhart, 2005), jednak późniejsze formy wykazywały już tendencję do zmniejszania się kinetyki czaszki i komplikacji morfologii koron zębowych (Lingham-Soliar i Nolf, 1989; Lingham-Soliar, 1988, 1995). Biorąc pod uwagę rozmiar, agresywność i niezwykłe przystosowania do drapieżnego stylu życia, bynajmniej ci więksi przedstawiciele Mosasauridae ( m.in. Mosasaurus hoffmanni, Tylosaurus proriger, Hainosaurus bernardi, Prognathodon saturator) były prawdopodobnie jednymi z najgroźniejszych morskich drapieżników w historii ziemi. Niektóre późne Mosasauridae wykazywały niezwykłe adaptacje żywieniowe jak na przykład przedstawiciele rodzajów Carinodens i Globidens miały kulisty kształt korony zęba przystosowany do gryzienia skorup mięczaków (Everhart, 2005; Poynter, 2011; Ross, 2006).
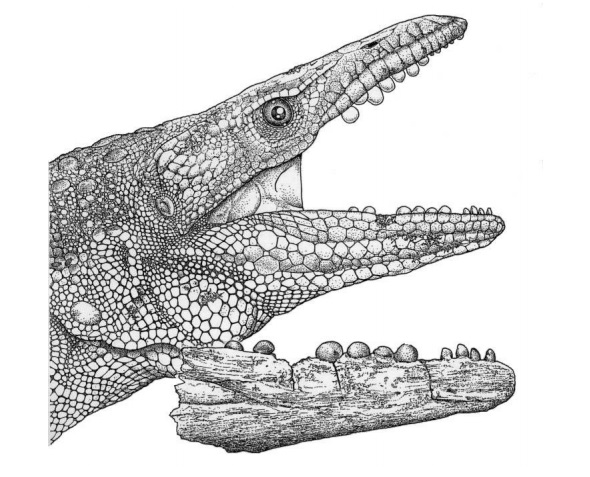
Globidens był przedstawicielem Mosasauridae, który ze względu na kulisty kształt korony zęba wykazywał tendencję do innego rodzaju drapieżnictwa, specjalizował się on mianowicie w pożywianiu się ofiarami o stosunkowo twardych łupinach (skorupiaki, amonity i być może mniejsze żółwie). Globidens mógł ukierunkować się w tego rodzaju diecie by wyeliminować potrzebę rywalizacji z innymi większymi przedstawicielami Mosasauridae zajmując najbardziej dogodną dla siebie niszę ekologiczną i po prostu wykorzystać obfitość zapasów żywnościowych w postaci wcześniej wymienionych ofiar.
Mosasauridae prowadziły bardzo agresywny styl życia, w swoich notatkach o formacji Kansas Benjamin Franklin Mudge (1877) opisał okaz w którym pięć kręgów grzbietowych zostało poważnie uszkodzonych tracąc wiele procesów kolczastych, egzemplarz przeżył ale kształt poszczególnych kości stracił naturalny kształt. Williston (1898) zauważył bardzo agresywne interakcje pomiędzy Mosasauridae o czym świadczyły liczne blizny i obrażenia jakich doznawali w ciągu życia. Williston donosi również, że obserwował patologie w dolnych szczękach i kręgach zwłaszcza ogonowych i kończynach, zdjęcie kończyn Platecarpus opublikowane przez Willistona pokazuje również fuzję w jedną masę po uszkodzeniu. W swojej książce ‘’Life of a Fossil Hunter’’ Sternberg (1909) wspomina o licznych patologiach kości Mosasauridae na które często natrafiali łowcy skamieniałości. Russel (1967) stwierdził, iż ciężkie obrażenia powodowane są u Mosasauridae przez osobniki tego samego gatunku, co wskazywało na gwałtowne konfrontacje pomiędzy nimi. Rothschild i Martin (1993), że niektóre z wyleczonych ran na ciałach Mosasauridae można by przypisać bardzo prymitywnej formie zalotów lub walk terytorialnych charakterystyka przypominające te jak u współczesnych jaszczurek, gdy jeden samiec próbuje złapać drugiego i odwrócić go na ‘’plecy’’. Według Rothschilda i Martina (1993) ząb, który utknął u podstawy lewej kości kwadratowej Mosasaurus condon jest interpretowany jako bezpośredni dowód śmiertelnego ataku osobnika tego samego gatunku (ten egzemplarz zawiera również inne ślady uszkodzeń). Znaleziona w 2012 roku próbka Mosasaurus missouriensis znana jest z wyleczonych nakłuć z zębów krewniaka.
Lingham-Soliar (2004) omówił paleopatologię i gojenie się obrażeń zaobserwowanych w dolnych szczękach Mosasauridae, w tym złamaną prawą egzemplarzu Platecarpus ictericus BMNH R 2834, który znajduje się w kolekcji British Museum of Natural History. Czaszka Clidastes propython PR 495 z kolekcji Field Museum of Natural History ma liczne częściowo wyleczone rany kłute na kościach czaszki. Sternberg Museum of Natural History w swoich zbiorach zawiera dwa eksponaty Tylosaurus kansasensis FHSM VP-3366 i VP-4516, które wyraźnie ukazują uszkodzenia w postaci złamanych żeber. Przykład Plioplatecarpus marshi MNHNP AC 9549-9775,9776 ukazuje uszkodzenie trzech kręgów kręgosłupa. Prawie kompletny szkielet Platecarpus tympaniticus FHSM VP-322 znaleziony w 1949 roku w hrabstwie Gov w stanie Kansas dostarcza dowodów na infekcję najprawdopodobniej spowodowaną ugryzieniem, która spowodowała połączenie dwóch kręgów bliżej ogona, a także pokazuje dwa inne miejsca fuzji kości na dolnej stronie ogona. Duża rana kłuta z oznaką gojenia jest widoczna na grzbietowej krawędzi nad prawą kością dolnej szczęki, a także drugie nakłucie w lewej kończynie przedniej co doprowadziło do połączenia dwóch paliczków jest dowodem, iż ten Platecarpus doznał w ciągu swojego życia wielu ran, otrzymanych od innych Mosasauridae w wyniku interakcji społecznych lub drapieżnictwa. Logicznym może być założenie, że duże Mosasauridae były jescze bardziej agresywne, pomimo mniejszej liczby próbek o dobrym stanie konserwacji różne patologie opisywano również dla wielkich okazów Mosasauridae z rodzajów : Tylosaurus, Mosasaurus i Prognathodon (Everhart, 2008; Schulp i współautorzy, 2006; Christiansen i Bonde, 2002; Schulp i współpracownicy, 2004; Bastiaans i współautorzy, 2014; Lingham-Soliar, 1998 , 2004). Niektóre Mosasauridae wykazują całkowicie odrębne patologie jak na przykład głęboką martwicę naczyniową bardzo często obecną u rodzajów: Platecarpus, Tylosaurus, Mosasaurus , Plioplatecarpus, Prognathodon, Hainosaurus i nieco mniej rozpowszechnioną u bardziej arktycznych Mosasauridae: Klidastes, Ectenosaurus, Globidens, Halisaurus i Plotosaurus (Bruce i współautorzy, 2007).

Grafika ukazująca agresywną interakcje pomiędzy dwoma przedstawicielami Mosasauridae: Tylosaurus pembinensis autor – Christopher Chavez.
Podobnie jak u współczesnych waranowatych Mosasauridae miały pierścienie twardówki (opisane przynajmniej dla przedstawicieli rodzajów Tylosaurus , Platecarpus , Clidastes , Mosasaurus i Prognathodon ; Yamashita i współautorzy, 2015; Russel, 1967), które podtrzymywały kształt rogówki. Przykład Mosasaurus hoffmanni, sugeruje, że miał on dość dobrą wizję ze względu na duże oczodoły, choć jego widzenie obuboczne było słabo rozwinięte jak u większości innych Mosasauridae (między innymi: 22° dla Plotosaurus bennisoni, 29° dla Mosasaurus missouriensis, 35° dla Phosphorosaurus ponpetelegans – Konisi i współpracownicy,2015). Ta cecha encefalizacji może wynikać z faktu, że Mosasaurus hoffmanni zamieszkiwał dwuwymiarowe siedlisko otwartego morza (reprezentowane przez niebo i powierzchnię wody) i polował na dużą zdobycz, która nie wymagała nadzwyczajnie precyzyjnej zdolności do jasnej oceny odległości, podczas gdy na przykład bardziej rozwiniętym widzeniem obuocznym charakteryzował się zamieszkujący środowisko trójwymiarowe płytkich mórz z dużą liczbą roślin i raf – Plioplatecarpus (Lingham-Soliar, 1995).
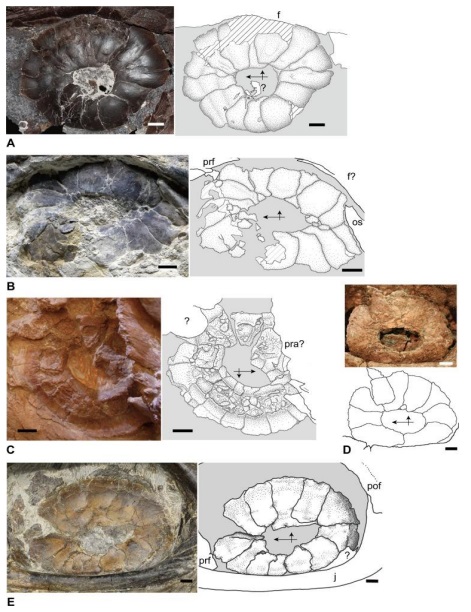
Grafika ukazująca materiał skalny zachowanych pierścieni twardówkowych poszczególnych przedstawicieli Mosasauridae: A. Tylosaurus proriger (FFHM 1997–10), B. Platecarpus tympaniticus (LACM 128319), C. Platecarpus tympaniticus (KU 1001), D. Clidastes propython (CFDC M74.10.06), E. Mosasaurus sp., cf. M. missouriensis (TMP 2012.010.0001).
Prawie na pewno Mosasauridae miały rozwidlony język, co zapewniało im rozwinięte poczucie chemorecepcji (chemoreceptory to receptory rozróżniające substancje chemiczne, do podstawowych chemoreceptorów zalicza się receptory wykrywające smak i zapach. Z chemorecepcją związane są również receptory wykrywające: tlen, dwutlenek węgla i Ph płynów ustrojowych). W oparciu o bardzo prawdopodobne położenie Mosasauridae w Platynota czyli grupą jaszczurek anguimorph – Schulp i współautorzy,2005 doszli do wniosku, że z pozycji związków filogenetycznych język u Mosasauridae najprawdopodobniej przypominał strukturalnie ten występujący u Helodermatidae oraz Lanthanotus borneensis (Waran głuchy), podczas gdy długi i wysoce wyspecjalizowany rozwidlony język taki jak u jaszczurek i węży uważany jest za mniej prawdopodobny. Oznacza to, iż rozwidlony język Mosasauridae był stosunkowo krótki a jego tylna część była powiększona, pokryta brodawkami i mukocytami (komórki kubkowe, rozmieszczone w błonie śluzowej), typowymi dla łuszczących się języków, co więcej język Mosasauridae prawdopodobnie nadal uczestniczył w przyjmowaniu pokarmu (funkcja ta jest zmniejszona u współczesnych waranowatych).
Nawias filogenetyczny prowadzi również do innego interesującego wniosku, Mosasauridae nie wykazują korelacji toksyczności dentystycznej i osteologicznej obserwowanej u toksycznych Platynota (grupa jaszczurek anguimorph, a zatem należy do rzędu Squamata klasy Reptilia. Odkąd został nazwany w 1839 roku, obejmuje kilka grup, w tym waranowate, węże, mozazaury i helodermatidy) takich jak członkowie rodziny Helodermatidae. Jednak obecnie wiadomo, że toksyczność występuje znacznie częściej u jaszczurek, niż wcześniej sądzono i że jadowite gruczoły żuchwowe rozwijały się nawet u przodków, choć w żadnym nie wykazywały korelacji osteologicznych ani dentystycznych (Fry i współpracownicy,2006). Tak więc jadowite gruczoły są podstawowym znakiem dla Mosasauridae jako przedstawicieli kładu Platynota, nie wiadomym jest jednak czy Mosasauridae zachowały trujące gruczoły czy je też utraciły podczas ewolucji. Ciekawym jest to, że pomimo rozwoju tekodontowego systemu dentystycznego, częstego narażania czaszki na uszkodzenia i całkowicie wodny styl życia, osteologiczne korelacje Mosasauridae wskazują, że ich zęby mogły być przysłonięte wargami, co być może wynikać z zachowania gruczołów żuchwowych zaangażowanych w proces trawienia.
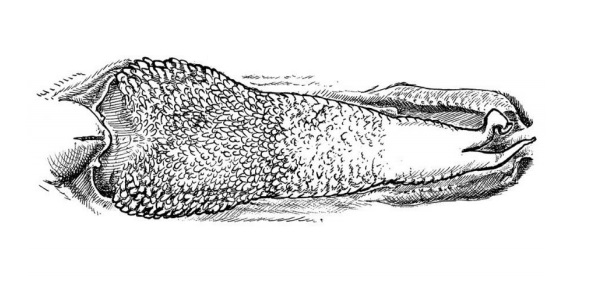
Szkic przedstawiający strukturę i wygląd języka Lanthanotus borneensis (Warana głuchego), prawdopodobnie bardzo podobną strukturą i formą charakteryzował się język u Mosasauridae.
Mózg u przedstawiciela Mosasauridae jakim był Platecarpus jako całość przypominał w strukturze ten u współczesnych waranowatych. Płaty węchowe są stosunkowo małe, co nie jest rzadkością w przypadku wtórnych kręgowców morskich, aczkolwiek pomocnicza opuszka węchowa jest dobrze rozwinięta, co wspiera użycie języka do chemorecepcji (Schulp i współpracownicy,2005). Płaty wzrokowe były dość duże, co najprawdopodobniej wskazuje, że Platecarpus skupiał się na widzeniu podczas polowania. Jak większość zwierząt Mosasauridae miały dwanaście par nerwów czaszkowych – nerwy czaszkowe III I IV były odpowiedzialne za ruchy gałek ocznych, a nerw czaszkowy VIII był odpowiedzialny za słyszenie i równowagę. Endocast (wewnętrzny odlew pustego obiektu, często odnoszący się do sklepienia czaszki w badaniach rozwoju mózgu u ludzi i innych organizmów) ukazuje wysoki poziom rozwoju półkolistych kanałów ucha wewnętrznego (Cuthbertson i współautorzy, 2015).
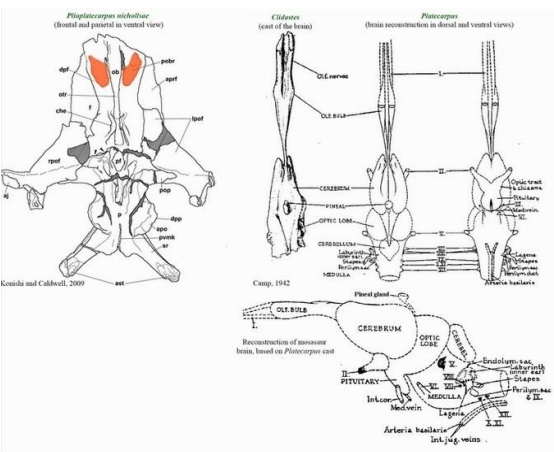
Grafika ukazująca kształt i ogólna lokalizację nerwów czaszkowych. Na przykładzie mózgu Platecarups widzimy, że płat węchowy znajdował się wzdłuż dolnej części czołowej, między gałkami ocznymi – był on stosunkowo niewielki co nie jest niczym niezwykłym u zwierząt morskich. Nerw wzrokowy był dobrze rozwinięty co sugeruje dobrą wizję w słabo oświetlonych lub głębszych wodach i wskazuje, że Mosasauridae ten był łowcą opierającym się na wizji. Ponieważ jednak oczy znajdowały się z boku łba nie wykazywały widzenia stereoskopowego. Podobnie jak u większości zwierząt mózgi Mosasauridae miały dwanaście par nerwów czaszkowych. I- nerw węchowy, II – nerw wzrokowy, V – nerw czuciowo-ruchowy, VI – nerw ten ma charakter ruchowy, unerwia mięsień prosty gałki ocznej, VII – nerw czaszkowy wchodzący w skład obwodowego układu nerwowego. Jest nerwem drugiego łuku skrzelowego. Nerw ma charakter mieszany, ale przeważają w nim włókna ruchowe. Zawiera także włókna czuciowe i przywspółczulne – wydzielnicze, VIII – nerw akustyczny, IX – nerw językowo-gardłowy, X – nerw błędny najdłuższy z nerwów czaszkowych, jest nerwem mieszanym, prowadzi włókna czuciowe, ruchowe i przywspółczulne, XI – nerw dodatkowy ma charakter ruchowy, a jego włókna mają początek w dwóch jądrach ruchowych: jądrze czaszkowym nerwu dodatkowego oraz jądrze rdzeniowym nerwu dodatkowego, XII – nerw podjęzykowy ma charakter ruchowy.
Potencjalne rozmiary największych przedstawicieli:
Mosasaurus hoffmanni / Mosasaurus lemonnieri
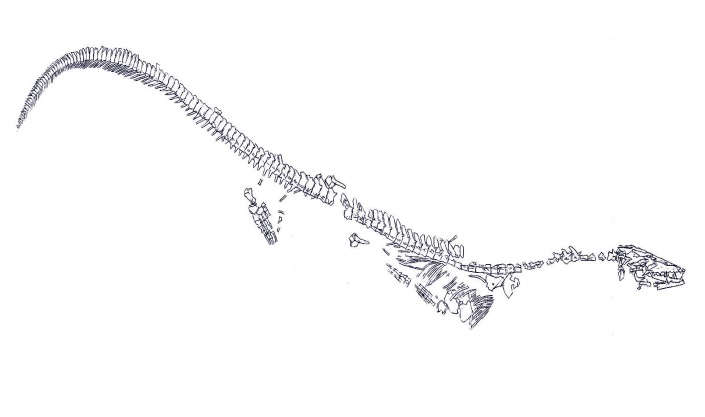
Szkic ukazujący zachowane elementy szkieletowe Mosasaurus lemonnieri IRSNB 3189.
Jeżeli chodzi o kwestię rozmiarową Mosasaurus hoffmanni nie będę się tutaj nadmiernie rozpisywał, gdyż oszacowanie wielkości tego gatunku jest szczegółowo przeanalizowane w umieszczonym przeze mnie artykule na witrynie DinoAnimals pod linkiem: https://dinoanimals.pl/blogi/czastytanow/mosasaurus-hoffmanni/. – *jedyne co chciałbym skorygować to kwestia uzyskanej masy określonej dla egzemplarzu CCMGE 10/2469, którą przeszacowałem, a sama kwestia wymaga ponownego rozpatrzenia. Kontynuując temat – Mosasaurus lemonnieri o numerze katalogowym IRSNB 3189 jest jednym z najlepiej zachowanych przedstawicieli Mosasauridae, gdzie w skład jego materiału kopalnego wchodzi przynajmniej 135 kręgów, mianowicie: 7 kręgów szyjnych, co najmniej 40 kręgów grzbietowych, 13 kręgów krzyżowych, 25 pośrednich kręgów ogonowych oraz około 50 końcowych kręgów ogonowych (Street,2016). Ogon u tego okazu jest proporcjonalnie nieco krótszy i zajmuje około 110% długości części przed kręgowej kręgosłupa i około 45% całkowitej długości zwierzęcia (Dollo, 1892). Oszacowanie całkowitej długości ciała dla tego dość kompletnego egzemplarzu Mosasaurus lemonnieri IRSNB 3119 z dolną szczęką osiągającą 1,14 metra i stosunku długości żuchwy do całkowitego rozmiaru ciała wynoszącego 1:11 (Dollo,1892) wynosiła by~ 12,54 metra.
Tylosaurus proriger / Tylosaurus pembinensis

Rekonstrukcja szkieletowa Tylosaurus proriger KUVP 5033 ‘‘Bunker‘‘, która jest elementem wystawowym Rocky Mountain Dinosaur Resource Center in Woodland Park, Colorado.
Północnoamerykańskie Tylosaurus są znane z dużej liczby stosunkowo kompletnych szkieletów, co czyni je najlepiej zbadanymi dużymi przedstawicielami Mosasauridae. Szkielet Tylosaurus składa się z 29-30 kręgów grzbietowych, 6-7 kręgów krzyżowych, 33-34 kręgi pośrednie ogonowe oraz 56-58 końcowych kręgów ogonowych, chociaż Russel,1967 podaje, iż liczba końcowych kręgów ogonowych u Tylosaurus proriger może osiągać 78. Czaszka jest stosunkowo duża a jej stosunek do całkowitej długości ciała wynosi około 1:7,15 (Everhart,2002), ogon natomiast zajmował 50 –55 % całkowitej długości (Russel,1967). Jeden z największych okazów pochodzący z wczesnego Kampanu skatalogowany jako KUVP 5033 znany również pod ksywą ‘’Bunker mosasaurus’’ ma czaszkę o długości ~1,83 metra (Konishi i współpracownicy,2018). W oparciu o oczekiwany rozmiar czaszki całkowita długość KUVP 5033 wynosiła by 13,08 metra. Everhart in współautorzy,2016 wskazali, że długość tego reprezentanta wynosi ~15,8 metra, aczkolwiek autorzy później stwierdzili, iż ta liczba była oparta na nieaktualnej rekonstrukcji sugerującej obecność krążków międzykręgowych dłuższych niż 1,3 cm i oczekiwanej liczby 140 kręgów. Konishi i współpracownicy,2018 wskazują długość KUVP 5033 wynoszącą prawie ~13 metrów. Niektóre inne wzorce materiału kopalnego Tylosaurus proriger z Kansas takie jak kręgi VP-642 i fragment uzębienia VP-2496 prawdopodobnie należeć mogą do osobników o podobnych rozmiarach do KUVP 5033. Everhart i współpracownicy,2016 informują także o skamieniałościach Tylosaurus porównywalnych rozmiarów do KUVP 5033 z Teksasu i Manitoby, a także o istnieniu być może jeszcze większych, ale słabo udokumentowanych okazów, które mogły osiągać ~14 metrów długości.
Dużym gatunkiem był także Tylosaurus pembinensis skatalogowany jako MT2 (holotyp), który pod względem wielkości porównywalny jest z największymi okazami Tylosaurus proriger takimi jak KUVP 5033. Niestety bardziej precyzyjne oszacowanie rozmiaru jest problematyczna ze względu na fakt, iż okaz wydaje się być mieszaniną dwóch różnych osobników z 137 – 138 kręgów MT 2 około 120 należy bezpośrednio do Bruce’a, a 15-20 należy do innego, znacznie mniejszego egzemplarzu (Lindgren i Siverson, 2002). Scott Hartman badając skamieniałe szczątki Tylosaurus pembinensis i dokonując rekonstrukcji graficznej szkieletu, gdzie deklarowana długość czaszki wynosi ~1,8 metra co sugerowało by wymiar zwierzęcia wynoszący 12,87 metra.
Hainosaurus bernardi

Rekonstrukcja szkieletowa Hainosaurus bernardi IRScNB R23 znajdująca się w Royal Belgian Institute of Natural Sciences.
Według wczesnych przedstawień Hainosaurus miał znacznie większą liczbę kręgów niż typowi przedstawiciele rodzaju Tylosaurus. Jeżeli ogon stanowił tę sama cześć całkowitej długości przedkręgowej części kręgosłupa co u Tylosaurus wówczas całkowita długość największego zaznanego okazu Hainosaurus bernardi o numerze katalogowym IRScNB R23 wynosiła by ~15 metrów (Lingham-Soliar, 1998). Holotyp Hainosaurus bernardi IRScNB R23 składa się z 41 kręgów szyjno – grzbietowych, 35 krzyżowych i / lub pośrednich kręgów ogonowych oraz 12 fragmentów, które mogą być końcowymi kręgami ogonowymi. Natomiast inny egzemplarz Hainosaurus bernardi IRScNB 3672 zawiera 38 kręgów szyjno – grzbietowych, 24 krzyżowych i / lub pośrednich kręgów ogonowych oraz 39 końcowych kręgów ogonowych, a także niewielką liczbę niezidentyfikowanych kręgów. Jeżeli założymy, że IRScNB 3672 zachowuje mniej lub bardziej kompletną serię kręgów ogonowych a kolumna kręgowa IRScNB R23 jest mniej więcej zakończona przed końcem serii pośrednich kręgów ogonowych, wówczas szkielet Hainosaurus bernardi składał by się z 41 kręgów szyjno- grzbietowych, 35 krzyżowych i /lub pośrednich kręgów ogonowych oraz 39 końcowych kręgów ogonowych. Całkowita liczba 115 kręgów będzie nieco mniejsza niż u blisko spokrewnionego Tylosaurus, co może wskazywać na niekompletność kręgosłupa u obu. Przedstawiciele rodzaju Tylosaurus mają zwykle 56- 58 końcowych kręgów ogonowych (Jimenez-Huidobro i Caldwell, 2015) co stanowi znacznie większą ilość niż 39 u IRScNB 3672, natomiast całkowita liczba kręgów krzyżowych i pośrednich kręgów ogonowych u Tylosaurus jest nieco mniejsza.
Zatem ogon Hainosaurus bernardi mógł być prawdopodobnie dłuższy niż sugeruje to złożona rekonstrukcja oparta na znanych pozostałościach kręgosłupa. Biorąc pod uwagę fakt, że całkowita długość pozostałej części kręgosłupa IRScNB R23 wynosiła by 8,1 metra Lindgren,2005 oszacował całkowitą długość kręgosłupa IRScNB R23 na 10,6 metra dodając brakujący końcowy odcinek ogona do rekonstrukcji od Tylosaurus. Wraz z czaszką o długości ~1,6 metra i jej współczynniku do całkowitej długość ciała 1:7,63 – IRScNB R23 osiągał by ~12,2 metra (Lindgren,2005), choć te szacunki Lindgrena należy jednak uznać za zachowawcze, ponieważ nie uwzględniają różnic w liczbie i proporcjach kręgów ogonowych u Tylosaurus proriger i Hainosaurus bernardi (pośrednie kręgi ogonowe IRScNB 3672 są porównywalne pod względem wielkości z holotypem Tylosaurus pembinensis ; Lindgren, 2005). Należy jednak zauważyć, że rekonstrukcja Hainosaurus bernardi na podstawie rekonstrukcji Tylosaurus pembinensis od Scotta Hartmana, zakłada mniej więcej taką samą całkowitą długość. Wskazana długość kręgosłupa IRScNB R23 nie uwzględnia żadnej przestrzeni międzykręgowej. Jeśli założymy, że IRScNB R23 miał taką samą przestrzeń międzykręgową, jak duży okaz Tylosaurus proriger (KUVP 5033), wówczas jego całkowita długość w ciągu życia byłaby o 38 cm dłuższa wynosząc 12,58 metra przy najbardziej zachowawczej liczbie odcinka szyjnego, grzbietowego, krzyżowego i pośrednich kręgów ogonowych. Gdyby Hainosaurus bernardi miał taką samą liczbę krzyżowych i pośrednich kręgów ogonowych jak Tylosaurus wówczas całkowita długość IRScNB R23, biorąc pod uwagę długość przestrzeni międzykręgowej będzie dłuższa o 42 cm wynosząc 12,62 metra. W obszarze końcowych kręgów ogonowych odległość między kręgami może być mniejsza, a istniejące obszary kostnienia stawów kręgów nie pozwalające na uwzględnienie długości przestrzeni międzykręgowej w tej części kręgosłupa (Rothschild i Everhart, 2015). Hainosaurus bernardi był prawdopodobnie cięższy niż typowi przedstawiciele rodzaju Tylosaurus o porównywalnej długości, , ponieważ dzięki zachowawczej rekonstrukcji miałby stosunkowo dłuższe ciało i krótszy ogon.
Prognathodon saturator / Prognathodon currii

Szkic materiału skalnego holotypu Prognathodon saturator NHMM 1998141 ukazujący zachowaną czaszkę oraz kręgów szyjno – grzbietowych.
Czaszka Prognathodon currii (HUJ. OR 100) ma długość żuchwy 149,7 cm i maksymalną długość górnej szczęki 142,2 cm (Christiansen i Bonde, 2002). Nieco większą niż czaszka holotypu Prognathodon saturator (NHMM 1998141) z 140 cm długością (Schulp i współautorzy,2016) charakteryzuje się ten sam gatunek o oznaczeniu IRScNB R24, gdzie dolna żuchwa jest o około 19,3 % dłuższa niż u holotypu (1,67 metra), niestety materiał pozaczaszkowy nie pozwala nam oszacować długości zwierzęcia. Sprawa ma się nieco bardziej optymistycznie, jeżeli chodzi o Prognathodon overtoni, które są dość dobrze zachowane. Prognathodon overtoni (TMP 2007.034.0001 i TMP 2002.400.0001) pozwalają na oszacowania wielkości, kolumna kręgowa Prognathodon overtoni składa się z 7 kręgów szyjnych, 33 kręgów grzbietowych, 11-12 kręgów krzyżowych, 22 kręgów pośrednich ogonowych i 37 końcowych kręgów ogonowych (chociaż nie wykluczona jest liczba 45, ponieważ liczba kręgów u Prognathodon overtoni zasadniczo odpowiada liczbie Clidastes liodontus). Całkowita długość TMP 2007.034.0001 wraz z czaszką wynosiła by ~5,9 metra, czaszka Prognathodon overtoni jest stosunkowo największa spośród wszystkich przedstawicieli podrodziny Mosasauridae o niezawodnie znanej proporcji stanowiącej około 15% całkowitej długości zwierzęcia. Konishi i współautorzy, 2011 podaje stosunek długości żuchwy do całkowitej długości ciała w opcji 1:7 (według pomiarów szkicu TMP 2007.034.0001). Stosując te zależność całkowita długość holotypu Prognathodon currii HUJ.OR 100 wynosiła ~ 10,48 metra, holotyp Prognathodon saturator ~9,8 metra, a egzemplarz IRScNB R24 ~11,69 metra, być może nieco więcej w zależności od długości przestrzeni międzykręgowej. Tak więc najwięksi przedstawiciele rodzaju Prognathodon były krótsze niż największe znane Mosasauridae. Jednak szacunki te niekoniecznie muszą być poprawne, ponieważ zgodnie z analizą filogenetyczną Simões i współpracowników, 2017 gatunki te niekoniecznie są ze sobą ściśle powiązane i mogą wykazywać różnice w liczbie i proporcjach kręgów.
Mosasaurus beaugei
Spośród stosunkowo kompletnych okazów czaszek Bardet i współautorzy,2004 wspominają o skatalogowanej jako OCP-DEK / GE 83, której długość wynosi 1,1 metra na podstawie zestawienia z Mosasaurus hoffmanni (stosunek wynosi 1:8,91) całkowita długość próbki OCP-DEK/ GE 83 można oszacować na 9,8 metra długości. Choć w tym miejscu należy zaznaczyć, iż znane są korony dentystyczne Mosasaurus beaugei o wysokości co najmniej 7,75 cm (korony zębów OCP-DEK / GE 83 mają 5 cm wysokości) i dość spory kawałek szczeki o długości 89 cm co może wskazywać na jeszcze większy rozmiar tego gatunku.
Kaikaifilu hervei
Kaikaifilu znany jest tylko z fragmentów czaszki, umożliwiających jednak rekonstrukcję całkowitej długości na około 1,1-1,2 metra. Aby ocenić pełną długość zwierzęcia można zastosować proporcję ciała innych przedstawicieli rodziny Tylosaurinae. U Hainosaurus bernardi stosunek długości czaszki do całkowitej długości ciała wynosi około 1:7,63 (Russel, 1967) co oznaczało by całkowitą długość w przedziale 8,39 – 9,15 metra, u Tylosaurus natomiast ten współczynnik wynosi 1:7,15 co dawało by nieco niższe wartości od 7,86 do 8,58 metra, co ogólnie prowadzi do szacunków całkowitej długości mniejszych niż 10 metrów wskazanych przez Otero i współpracowników,2016.
*Opublikowany artykuł może być wykorzystywany wyłącznie na własny użytek edukacyjny, nie może stanowić źródła informacyjnego dla innych blogów, wykluczonym jest także udostępnianie ani publikowanie go bez zgody autora.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Poprzedni artykuł:
Transakcje materiałem kopalnym i problematyka z nimi związana Następny artykuł:
Prognathodon
Transakcje materiałem kopalnym i problematyka z nimi związana Następny artykuł:
Prognathodon