
71 milionów lat temu lądami władały dinozaury, podobnie jak współcześnie większość planety pokrywają morza. Olbrzymie oceany to królestwa przemocy w głębinach jest bardziej niebezpiecznie niż na lądach, w tym brutalnym świecie ”narodził się” jeden z największych drapieżników morskich w dziejach naszej planety – Mosasaurus hoffmanni najgroźniejsze stworzenie w najbardziej okrutnym oceanie wszechczasów.

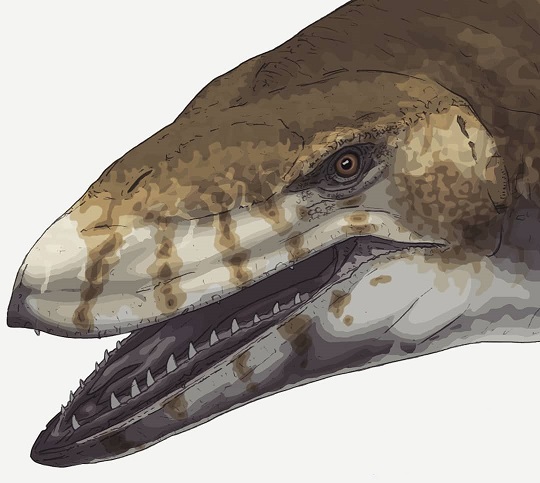
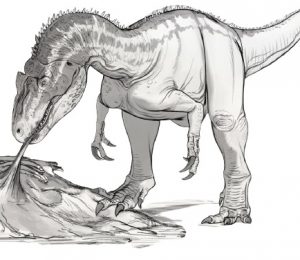
Model przedstawiający potencjalny wygląd za życia Mosasaurus hoffmanni autorstwa Goro Furuty.
Systematyka:
Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce
Gromada: zauropsydy – Podgromada: diapsydy – Nadrząd: lepidozaury
Rząd: łuskonośne – Rodzina: mozazaury – Rodzaj: Mosasaurus – Gatunek: M.hoffmanni
Czas i miejsce występowania:
Gatunek Mosasaurus hoffmanni (ok. 71 -66 mln lat temu, późna kreda – mastrycht) był kosmopolityczny tereny jego występowania obejmowały : w Belgię ( Jagt i współpracownicy, 2006 ), Jordanii ( Kaddumi, 2006 ), Rosję (Grigoriev, 2014 ), Turcję (Bardet i Tunoglu, 2002 ), Stany Zjednoczone ( Gallagher i współpracownicy, 2005 oraz Kiernan, 2002 ), Danie ( Lindgren i Jagt, 2005 ), Bułgarię ( Godefroit i Motchurova-Dekova, 2010 ), Polskę (Michalski i współpracownicy, 2003 ), Włochy ( Palci i współpracownicy, 2014 ), oraz Angole ( Mateus i współpracownicy, 2012).
Znaczenie nazwy rodzajowej i epitetu gatunkowego:
Nazwa rodzaju Mosasaurus związana jest z rzeką Mosa w obrzeżach której znajdowała się kamieniołom w którym po raz pierwszy odnaleziono pierwszy materiał skalny Mosasaurus hoffmanni oraz greckiego ”saurus”- jaszczurka i w wolnym tłumaczeniu można ją interpretować jako jaszczura znad rzeki Mosa. Epitet gatunkowy hoffmanni honoruje Johanna Leonarda Hoffmanna. Nazwa została nadana w 1829 roku przez Gideona Montella.
Szacunek wielkości i masy:
Na chwilę obecną za największego przedstawiciela gatunku Mosasaurus hoffmanni uznaje się okaz skatalogowany jako CCMGE 10/2469 którego długość szacuje się na 15,24 metra a masę na 18869 kilogramów. Niektóre fragmentaryczne próbki wskazują, że Mosasaurus hoffmanni może osiągnąć większe rozmiary niż największe znane osobniki z dobrze zachowanymi żuchwami (NHMM 009002, CCMGE 10/2469 i ALMNH PV 988.0018), jednak ich oszacowania się nie podejmę , gdyż nie preferuje porównań bazujących na rozmiarach pojedynczych elementów, ponieważ są one szalenie spekulacyjne.
Odkrycie i klasyfikacja:
Pierwszą czaszkę należącą do Mosasaurus hoffmanni odkrył porucznik Jean Baptiste Druon w 1764 roku w wapiennym kamieniołomie w Monte San Pedro niedaleko Maastricht miasta położonego w południowej Holandii. W 1766 roku znalezisko zostało przeniesione do Teylers Museum, gdzie Martin van Marum ówczesny dyrektor wcześniej wspomnianego muzeum w roku 1874 opublikował swój opis dotyczący czaszki klasyfikując ją jako należącą do walenia lub ryby nieznanej jeszcze nauce. Ta kopia skatalogowana jako TM 7424 po dziś dzień jest częścią kolekcji muzeum.
Obecnie Mosasaurus camperi, Mosasaurus dekayi, Mosasaurus giganteus oraz Mosasaurus maximus uważane są za gatunki synonimiczne do Mosasaurus hoffmanni (Mulder,1999 oraz Street, 2016). Oprócz Mosasaurus hoffmanni niezależne gatunki Mosasaurus są rozpoznawane jako Mosasaurus missouriensis, Mosasaurus conodon, Mosasaurus lemonnieri, Mosasaurus beaugei (Street,2016). Chociaż współczesne badania filogenetyczne (Simões i współpracownicy,2017), a także starsze obserwacje (Russel,1967), nie potwierdzają umiejscowienia Mosasaurus conodon w obrębie tego rodzaju. Zamiast tego Mosasaurus hoffmanni jest bardzo blisko spokrewniony z Plotosaurus (Simões i współpracownicy, 2017), ponadto (Grigoriev,2014) sugeruje, iż Mosasaurus lemonnieri jest młodym osobnikiem Mosasaurus hoffmanni.
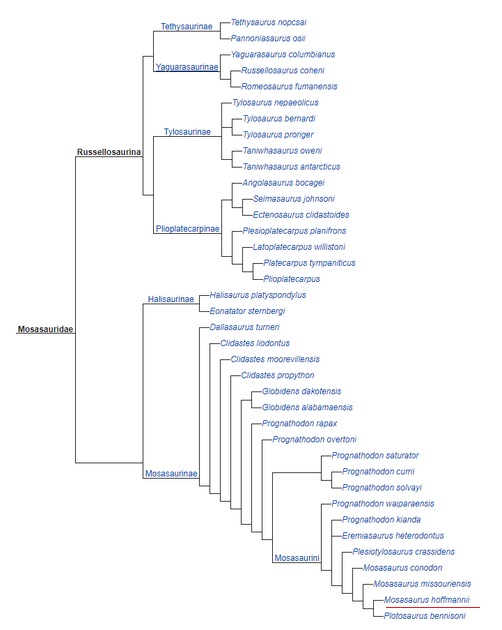
Kładogram z pozycją systematyczną Mosasaurus hoffmanni według Simoes i współpracownicy(2017).
Względnie duże oczodoły wskazują, że Mosasaurus hoffmanni miał dobrą wizję, chociaż jego widzenie obuoczne było słabo rozwinięte jak u większości innych mozazaurów (między innymi: 22° dla Plotosaurus bennisoni, 29° dla Mosasaurus missouriensis, 35° dla Phosphorosaurus ponpetelegans – Konisi i współpracownicy,2015). Ta cecha encefalizacji może wynikać z faktu, że Mosasaurus hoffmanni zamieszkiwał dwuwymiarowe siedlisko otwartego morza (reprezentowane przez niebo i powierzchnię wody) i polował na dużą zdobycz nie wymagała nadzwyczajnie precyzyjnej zdolności do jasnej oceny odległości, podczas gdy na przykład bardziej rozwiniętym widzeniem obuocznym charakteryzował się zamieszkujący środowisko trójwymiarowe płytkich mórz z dużą liczbą roślin i raf Plioplatecarpus (Lingham-Soliar, 1995). Badania izotopu węgla w skamieniałych szczątkach Mozazaurów sugeruje, iż więksi przedstawiciele rodziny nurkowały na większe głębokości niż ich mniejsi krewni (średnia głębokość ok 50 metrów – Lingham-Soliar i Nolf,1989). Sytuacja paleograficzna w innych miejscach w których odkryto Mosasaurus hoffmanni jest mniej zrozumiała. W Ameryce Północnej Mosasaurus maximus = Mosasaurus hoffmanni najwyraźniej zamieszkiwał zarówno płytkie jak i głębokie wody, zastępując równie duże Tylozaury (Russel, 1967; Kiernan, 2002). Co ciekawe w Europie duzi przedstawiciele podrodziny Tylosaurinae jak np. Hainosaurus bernardi współistniały z dużymi przedstawicielami podrodziny Mosasaurinae jak Mosasaurus hoffmanni oraz Prognathodon saturator. Konkurencyjny sukces europejskich tylozaurów w porównaniu z ich krewnymi z Ameryki Północnej był prawdopodobnie spowodowany ich większym stopniem rozwoju ewolucyjnego (Lingham-Soliar, 1995).
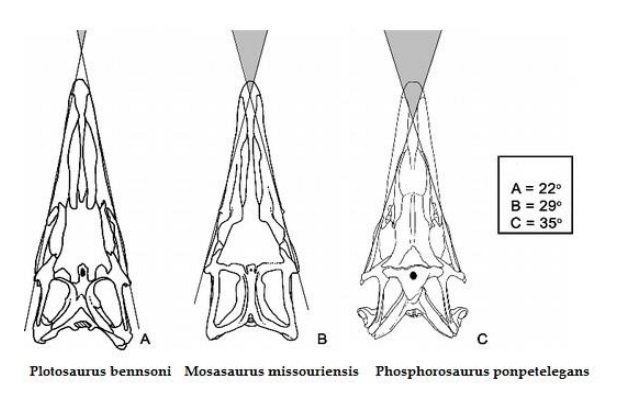
Rekonstrukcja widzenia obuocznego u wybranych mozazaurów.
Struktura ogona i kończyn wskazuje, że Mosasaurus hoffmanni był szybkim, ale niezbyt zwrotnym pływakiem (Russel, 1967). Stosunkowo niewielki rozmiar płetw i kształt tułowia (maksymalna wysokość klatki piersiowej jest w przybliżeniu równa jego maksymalnej szerokości) prawdopodobnie ograniczała jego mobilność podczas ostrych skrętów (Lingham-Soliar, 1995). Duże kości biodrowe Mosasaurus hoffmanni konkretnie okazu skatalogowanego jako TSMHN 11208 ma szereg obrażeń osteologicznych, których przyczyną prawdopodobnie była regularna rotacja kończyn, gdy Mozazaur obracał masywne ciało (Lingham-Soliar, 1995). Ogólnie rzecz biorąc Mosasaurus hoffmanni mógł mieć podobne adaptacje pływackie do blisko spokrewnionego Plotosaurus bennisoni (Russel, 1967; Lingham-Soliar, 1995; Lindgren i współpracownicy, 2011), który jest prawdopodobnie najbardziej wyspecjalizowanym pływakiem wśród wszystkich mozazaurów (Lindgren i współpracownicy,2011). Płetwa ogonowa Mosasaurus hoffmanni najprawdopodobniej przypominała kształt płetwy ogonowej Prognathodon (ERMNH HFV 197) ale ze względu na niedojrzałość ERMNH HFV 197 i większe ‘’zaawansowanie’’ ewolucyjne rodzaju Mosasaurus wydaje się prawdopodobne, że płetwa ogonowa dorosłego Mosasaurus hoffmanni miała by głębszą formę niż w przypadku ERMNH HFV 197 (korespondencja osobista z Lindgren). Żebra Mosasaurus hoffmanni są stosunkowo grubsze a kręgi masywniejsze niż u innych mozazaurów takich jak Hainosaurus i Plotosaurus (Lingham-Soliar, 1995).

Zrekonstruowany szkielet Mosasaurus hoffmanni NHMM 006696 w Muzeum Historii Naturalnej w Maastricht – Holandia. Model został zrekonstruowany przez Hansa Brinkerinka w drugiej połowie XX wieku. Ze względu na niewystarczającą wiedzę przedstawicieli rodzaju Mosasaurus w momencie jego powstania, rekonstrukcja zawiera wiele niedokładności, szczególnie w kręgosłupie, w tym jeden dodatkowy kręg szyjny, niewystarczająca liczba kręgów grzbietowych i krzyżowych (Schulp, korespondencja osobista).
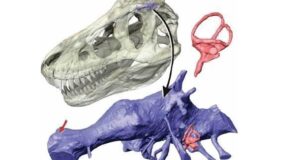
Adaptacja czaszki Mosasaurus hoffmanni jest nietypowa dla większości innych mozazaurów, kinetyzm czaszek wcześniejszych członków rodziny został prawie całkowicie utracony zachowane zostały tylko streptostylia i staw śródżuchwowy. Kluczową rolę w redukcji stref metakinetycznych i mezokinetycznych w czaszce Mosasaurus hoffmanni stanowiło ‘’zakładanie’’ się na siebie grzbietowej kości ciemieniowej z kością czołową.
Mosasaurus hoffmanni wykazuje najbardziej zaawansowaną morfologię zębów spośród wszystkich mozazaurów, oprócz dwóch ząbkowanych cariane, które dzielą koronę zęba na dwie boczne powierzchnie, każda korona ma kilka dodatkowych powierzchni tnących, których liczba zwykle odpowiada dwóm z przodu i pięciu z tyłu. Zęby szczęki i kości zębowej są powiększone w porównaniu z wieloma innymi mozazaurami, podczas gdy zęby przedmaxilla są stosunkowo małe. Korony dentystyczne Mosasaurus hoffmanni są zaprojektowane tak aby wytrzymywały duże naprężenia mechaniczne powstające w wyniku silnego zgryzu, a zatem nadawały się dobrze rozdzierania mięsa i miażdżenia kości (Massare, 1987; Street, 2016; Lingham-Soliar, 1995). Żeby na pterygoidzie (ich liczba sięga 8 z każdej strony) u Mosasaurus hoffmanni były znacznie zmniejszone co utrudniało mu karmienie mechanizmem zapadkowym, który jest charakterystyczny dla wielu innych mozazaurów. Mosasaurus hoffmanni jednak częściowo zrównoważał to streptostiliami (ruchome mocowanie kwadratowej kości), które pozwalały żuchwie poruszać się do przodu i do tyłu w większym zakresie, bardziej ruchomy staw żuchwowy pozwalał Mosasaurus hoffmanni do bardziej wyspecjalizowanych manipulacji takich jak choćby cięcie ofiary zębami czy stabilizowanie żuchwy podczas maksymalnej siły ugryzienia (to ostatni obserwuje się we współczesnych waranowatych jak choćby Varanus exanthematicus ; Lingham-Soliar i Nolf, 1989; Lingham-Soliar, 1995).
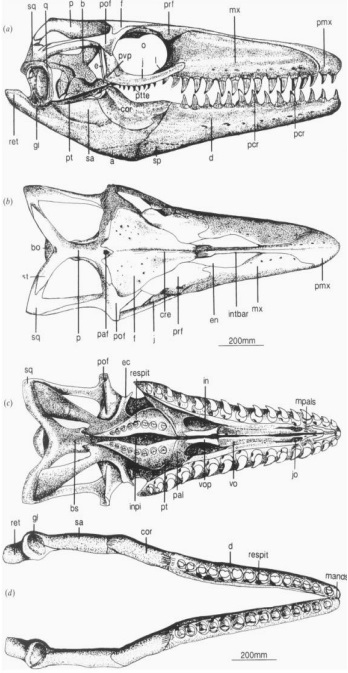
Rekonstrukcja czaszki Mosasaurus hoffmanni autorstwa Lingham-Soliar.
Jak pokazują odciski na kościach czaszek niektórych okazów Mosasaurus hoffmanni mięśnie ich mięśnie czaszkowe były bardziej zróżnicowane niż wielu innych mozazaurów i zapewniały większą kontrolę nad ruchami czaszki i żuchwy. Powierzchnia kości balszkowatej na przednie rozszerzenie mięśni żuchwy ( Musculus adductor mandibulae externus lub Musculus pseudotemporalis profundus ) morfologicznie przypominające te jakie występują u współczesnych jaszczurek Tupinambis teguixin lub Xenosaurus grandis, które mają dość dużą siłę ugryzienia jak na swój rozmiar. Powierzchnia koronoidu wyraźnie wskazuje na obecność dużego mięśnia przywodziciela zewnętrznego, co wskazuje na zdolność Mosasaurus hoffmanni do ściskania szczek z dużą siłą, mocowanie tego mięśnia głównie po wewnętrznej stronie żuchwy zapewniało skuteczne rozłożenie obciążeń po obu stronach żuchwy.
Ponadto inne mięśnie żuchwy (Musculus pseudotemporalis superficialis i Musculus pseudotemporalis profundus) wytwarzały silny moment obrotowy wokół własnej osi podłużnej i mogły prowadzić do obrotu przyśrodkowego dolnej szczęki i tworzenia obciążeń rozciągających po stronie grzbietowej osłabionego spojenia (osłabienie to wynika ze względu na fakt, iż naziemni przodkowie mozazaurów połykali swoje ofiary w całości jak współczesne węże) podczas ściskania szczęk. Mięśnie Musculus pterygoideus, które były przymocowane do bocznej powierzchni każdej strony żuchwy w trakcie procesu ewolucyjnego całkowicie wyeliminowały tę niestabilność. Mięśnie przywodziciela Mosasaurus hoffmanni działały również jako ‘’ oryginalne pasma architektoniczne’’ wzmacniając dolną szczękę (Lingham-Soliar, 1995). Na podstawie siły zgryzu Teju argentyńskiego (Tupinambis merianae) maksymalną siłę zgryzu metodą skalowania izometrycznego u Mosasaurus hoffmanni skatalogowanego jako CCMGE 10/2469 (PRM2546) o długości dolnej szczęki wynoszącej 171 cm można oszacować na 110893 niutonów lub 11307,93 kg/cm3. Wzrost mięśni odpowiedzialnych za otwieranie szczęk prawdopodobnie nie był zbyt znaczny w przypadku Mosasaurus hoffmanni ze względu na potencjalnie dużą masę żuchwy i umiarkowaną szerokość czaszki. Ścięgna przyczepione do koronoidów mogły działać jako dodatkowa siła ograniczająca przy maksymalnym otwarciu szczęk, co zwiększyło szybkość ich zamykania (Lingham-Soliar, 1995).
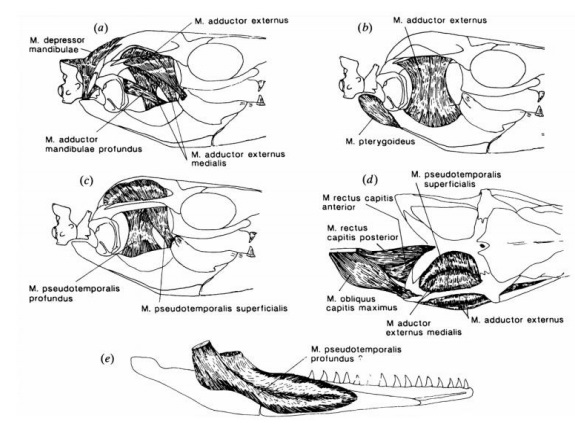
Rekonstrukcja mięśni czaszki Mosasaurus hoffmanni autorstwa Lingham-Soliar.
Według wskazań Mosasaurus hoffmanni był dominującym drapieżnikiem morskim, który polował głownie na stosunkowo dużą zdobycz przy powierzchni wody, jego głównym celem były inne mozazaury i plezjozaury pływające przy powierzchni w celu zaczerpnięcia powietrza. W tej sytuacji potencjalna ofiara była najbardziej narażona na atak, ponieważ znajdowała się w dobrze oświetlonej części wody i mogła być łatwo wykryta przez drapieżnika pływającego pod nim, a także nie była w wielu wypadkach odpowiednio szybko zareagować. Solidna budowa czaszki Mosasaurus hoffmanni doskonale nadawała się do wytrzymywania obciążeń wynikających z oporów dużej ofiary, podczas gdy duża siła ugryzienia i synchroniczne ruchy dolnej szczęki tam iż powrotem umożliwiały temu gatunkowi odgryzanie dużych kawałków mięsa od potencjalnej ofiary. Biorąc pod uwagę siłę czaszki Mosasaurus hoffmanni prawdopodobnie mógłby również potrząsnąć schwytaną ofiarą, a nawet obracać nią. Wielkość, masa i domniemana agresywność dorosłych osobników Mosasaurus hoffmanni sugerują, że prawdopodobnie mógłby pokonać prawie każde zwierzę morskie z późnej kredy.
Potencjalne spektrum ofiar Mosasaurus hoffmanni jest duże, w Ameryce Północnej gatunek ten współistniał z Elasmosauridae takimi jak Cimoliasaurus magnus (Baird, 1986), Aphrosaurus furlongi (Stock, 1939), Fresnosaurus drescheri (Ford, 2006), Hydrotherosaurus alexandrae (Welles, 1940 i Morenosa Ford, 2006) i innymi mozazaurami w tym Mosasaurus sp. (Mosasaurus Conodon i innych niezidentyfikowanych skamielin być może należących do Mosasauridae; Baird, 1986; Gallagher, 2005; Kiernan, 2002), Prognathodon sp. ( Prognathodon rapax i Prognathodon waiparaensis ; Baird, 1986; Lindgren, 2009), Plesiotylosaurus crassidens (Lindgren, 2009) i Plotosaurus bennisoni (Lindgren i współpracownicy, 2008), okazami z podrodziny Mosasaurinae: Plioplatecarpus sp. ( Plioplatecarpus depressus =? Plioplatecarpus marshi, Plioplatecarups primaevus ; Mulder, 1999; Kiernan, 2002; Tokaryk, 1993; Gallagher, 1993) oraz okazami z podrodziny Halisaurinae i Plioplatecarpinae (Baird, 1986), krokodylami takimi jak : Thoracosaurus neocesariensis i Hyposaurus rogersii (Baird, 1986; Gaffney i współpracownicy., 2006) oraz dużą liczbą gatunków żółwi morskich na przykład: Osteopygis spp., Peritresius ornatus , Catapleura ponderosa i Lytoloma jeanesii (Gallag, 1993; Hay, 1908; Cope, 1871).

Mozazaury współistniejące z gatunkiem Mosasaurus hoffmanni autorstwa paleosir (* nie należy się tutaj sugerować skalą wielkości poszczególnych gatunków tak jak to zobrazował wcześniej wymieniony autor, gdyż jest moim zdaniem ona błędna). Długość ciała gatunków Mosasaurus hoffmanni / lemonnieri jest zaniżona. Z określeniem proporcji Prognathodon saturator mam pewne obiekcje, ponieważ przedstawiciele rodzaju Prognathodon mają inną liczbę kręgów i być może zupełnie inne proporcje (Konishi i współpracownicy, 2011), więc długość największych Prognathodon może wahać się w przedziale od 11,69 metra do 12,74 metra. Gdy będę miał nieco więcej czasu postaram się stworzyć prawidłową skalę powyższych gatunków i zastąpić nią tę którą aktualnie widzicie.
Na wodach pokrywających część terytorium współczesnej Afryki i Europy Mosasaurus hoffmanni współistniał z innymi mozazaurami w tym Mosasaurus sp. (Mosasaurus lemonnieri, Mosasaurus
beaugei; Abdel-Gawad, 1986; Bardet, 2012; Lingham-Soliar, 2000) , Prognathodon sp. (Prognathodon solvayi, Prognathodon kianda, Prognathodon sectorius, Prognathodon currii, Prognathodon saturator, Prognathodon giganteus ; Dortangs i współpracownicy., 2002; Schulp i współpracownicy., 2008; Christiansen i Bonde, 2002; Jagt i współpracownicy., 2006; Bardet, 2012,2005; Bardet i współpracownicy., 2000; Lingham-Soliar i Nolf, 1989), Globidens phosphaticus (Polcyn i współpracownicy, 2010; Bardet, 2012), Carinodens sp. (Carinodens belgicus, Carinodens minalmamar ; Jagt i współpracownicy, 2006; Schulp i współpracownicy., 2009), Liodon sp. (Liodon anceps i Liodon sectorius (Kaddumi, 2006; Mulder i współpracownicy, 1998), Eremiasaurus heterodontus (LeBlanc i współpracownicy, 2012) i Igdamanosaurus aegyptiacus (Bardet, 2012) okazami z podrodziny Mosasaurinae , Halisaurus sp. (Halisaurus ortliebi i Halisaurus arambourgi; Jagt i współpracownicy., 2006) i Phosphorosaurus ortliebi (Konishi i in., 2015) z podrodziny Halisaurinae : Plioplatecarpus sp. (Plioplatecarpus houzeaui, Plioplatecarpus marshi : Jagt i współpracownicy, 2006, Bardet, 2012; Mulder i współpracownicy, 1998) z podrodziny Plioplatecarpinae , Hainosaurus bernardi (Jagt i współpracownicy, 2006) i Goronyosaurus nigeriensis (Azzaroli i współpracownicy, 1972). Żółwiami morskimi: Gigantatypus salahi (Kaddumi, 2006), Allopleuron hofmanni (Lingham-Soliar, 1995; Mulder i współpracownicy, 1998), Glyptochelone suyckerbuyki , Plaechelone emarginata , Calcarichelys sp., Protostega sp. i Euclastes sp. (Mateus i współpracownicy, 2012), krokodylami: Dyrosaurus phosphaticus, Rhabdognathus sp., Phosphatosaurus sp. i Thoracosaurus sp. (Gemmellaro, 1921; Jouve, 2007; Mulder i współpracownicy, 1998), plezjozaurami: z rodzin Polycotylidae (Kaddumi, 2006) i Elasmosauridae (Jagt i współpracownicy, 2006; Mulder i współpracownicy, 1998; Mulder i współpracownicy, 2000) takie jak: Tuarangisaurus sp. (Mateus i współpracownicy, 2012), Goniosaurus binkhorsti i Zarafasaura oceanis (Vincentet, 2011).Większość tych gadów morskich była potencjalną ofiarą Mosasaurus hoffmanni, raczej wąska powierzchnia narządu Jacobsona sugeruje, że zmysł węchu prawdopodobnie nie odgrywał tak istotnej roli w życiu Mosasaurus hoffmanni , jak u niektórych innych mozazaurów, ta cecha może być związana ze stylem życia pelagicznego i polowaniem w pobliżu powierzchni wody, ponieważ w tym przypadku wzrok odgrywa ważniejszą rolę w wykrywaniu potencjalnej ofiary niż zmysł węchu (Lingham-Soliar, 1995). Jednocześnie padlina nie była znacząca częścią diety Mosasaurus hoffmanni, ponieważ stosunkowo słabo rozwinięty zmysł węchu oznaczał mały promień wykrywania martwych tuszy i nie przyczyniał się do konkurencyjnego sukcesu w niszy ekologicznej, jeżeli chodzi o kwestię padlinożerności w porównaniu z innymi mozazaurami i rekinami.

Grafika ukazująca Mosasaurus hoffmanni podczas polowania na rodzaj wymarłej ryby Xiphactinus audax, oraz współistniejące z nim gatunki: 1. Pterandon stanbergi – 2. Hesperornis regalis – 3. Enchodus petrosus – 4. Cimolichthys nepaholica – 5. Squalicorax bassanii – 6. Protostega gigas
Niestety nie ma próbek koprolitów czy zawartości żołądkowych przypisywanych gatunkowi Mosasaurus hoffmanni, istnieją jednak inne ‘’paleo dowody’’ na drapieżna aktywność tego drapieżnika morskiego. Pancerz wymarłego żółwia morskiego Allopleuron hofmanni (od 2 do 2,5 metra długości) nosi ślady zębów dużego mozazaura prawdopodobnie Mosasaurus hoffmanni (Lingham-Soliar, 1995) lub Prognathodon saturator (Dortangs, 2002). Dwa młode osobniki Mosasaurus hoffmanni (IRSNB R25 i IRSNB R27 o długościach czaszek wynoszących odpowiednio 60 cm i 80 cm) wykazują poważne uszkodzenia żuchwy, prawdopodobnie spowodowane naprężeniami mechanicznymi podczas atakowania stosunkowo dużej i silnej ofiary (Lingham-Soliar i Nolf, 1989; Lingham-Soliar, 2004). W przypadku większych okazów Mosasaurus hoffmanni takie patologie nie zostały jeszcze opisane, co najprawdopodobniej wynika z faktu, że dorosły Mosasaurus hoffmanni był jednym z największych zwierząt morskich późnej kredy i z trudem mógł napotkać opór, który mógłby zniszczyć ich masywne szczęki. Niezidentyfikowany okaz TMP 83.126.1, najwyraźniej na krótko przed śmiercią zjadł mosasauridae z rodziny Plioplatecarpinae niezidentyfikowane zwierzę (Bell i Barnes, 2007). Jeden mały przedstawiciel gatunku Mosasaurus missouriensis (TMP 2008.036.0001) z czaszką o długości około 66 cm w rejonie żołądka zawierał resztki kości ryby kostnej o długości nieco większej niż metr z rzędu Aulopiformes (Konishi i współpracownicy, 2014).
Wiele próbek Mosasaurus hoffmanni w tym czaszka holotypu, nosi ślady urazów śródmiąższowych spowodowanych przez krewnych (Street i Caldwell, 2016). Kanibalizm był prawdopodobnie bardzo
powszechny u Mosasaurus hoffmanni i nawet duże dorosłe osobniki doświadczyły aktów wewnątrzgatunkowej agresji ze strony krewnych, o czym świadczy na przykład ugryziona kwadratowa kość i lewa część dolnej szczęki próbki NHMM 006696 (Schulp i współpracownicy,2006), ponadto istnieją pewne pośrednie dowody agresywnych interakcji międzygatunkowych między Mosasaurus hoffmanni innymi gigantycznymi mozazaurami. Młody okaz skatalogowany jako IRSNB R12 z dolną szczęką o długości ok. 1,1 metra ‘’pokazuje’’ pękniecie czaszki i przemieszczenie się pnia mózgowego, a także zwichnięcie koronyda i kość łukowej po prawej stronie dolnej szczęki. Jedna z interpretacji takich poważnych obrażeń jest uderzenie przez dużego tylozaura Hainosaurus bernardi ( Lingham-Soliar, 1998). Czaszka innego dużego gatunku Prognathodon currii ma dziurę w kości czołowej, biorąc pod uwagę kształt uszkodzenia, głównym podejrzanym drapieżnikiem zdolnym do ataku jest Mosasaurus hoffmanni (Christiansen i Bonde, 2002).
Grafika ukazująca agresywna interakcję pomiędzy Mosasaurus hoffmanni.
Obecnie uważa się, że najwiarygodniejszą metodą oceny wielkości Mosasaurus hoffmanni jest skala z żuchwy, ale nawet bez oceny całkowitej długości żuchwy można powiedzieć, że był to jeden z największych mozazaurów. Russel (1967) wskazał, że żuchwa zajmuje około 10% całkowitej długości Mosasaurus maximus = Mosasaurus hoffmanni. Następnie ten stosunek został wykorzystany przez różnych autorów (np. Lingham-Soliar, 1995; Grigoriev, 2014) w celu oszacowania wielkości niektórych próbek Mosasaurus hoffmanni, jednak w tej chwili nie jest do końca jasne, jak prawdziwe są te proporcje, ponieważ żadna z próbek Mosasaurus maximus = Mosasaurus hoffmanni nie była znana w momencie publikacji artykułu przez Russel (1967) co uniemożliwiało wiarygodne oszacowanie wielkości i proporcji zwierzęcia (np. próbek TMM 313 i NJSM 11053). Wśród innych przedstawicieli podrodziny Mosasauridae, z których znane są stosunkowo kompletne szkielety, Plotosaurus bennisoni ma stosunek długości dolnej szczęki do całkowitej długości w zakresie 1: 9,5 (rekonstrukcja kompozytowa według Lindgren i współpracowników, 2007) lub około 1: 8,5 (okaz skatalogowany jako UCMP 32778 według Lindgren i współpracownicy, 2008), Eremiasaurus heterodontus około 1: 8,6 (LeBlanc i współpracownicy, 2012), Clidastes liodontus około 1: 8,3 (Russel, 1967), Prognathodon overtoni około 1: 6,7 (Konishi i współpracownicy, 2011), Mosasaurus lemonnieri – 1:11 (Dollo, 1892), Mosasaurus missouriensis około 1: 7,2.
Wykorzystanie proporcji bliskich krewnych do oszacowania wielkości Mosasaurus hoffmanni jest skomplikowane przez fakt, że liczba kręgów u przedstawicieli rodzaju może wynosić od 128 u ( Mosasaurus conodon SDSM 452; 7 szyjnych, 38 grzbietowych, 8 krzyżowych, 21 pośrednich ogonowych i około 54 końcowych kręgów ogonowych – Russel, 1967; Ikejiri i Lucas, 2014) do 130 u (Mosasaurus missouriensis TMP 2012.010.0001; 7 szyjnych, 33 grzbietowych, 11 krzyżowych, 24 pośrednich ogonowych i 55 końcowych kręgów ogonowych; Street,2016) i co najmniej 135 u (Mosasaurus lemonnieri IRSNB 3119; 7 szyjnych, co najmniej 40 grzbietowych, 13 krzyżowych, 25 pośredniego ogonowych i około 50 końcowych kręgów ogonowych; Street,2016). Jednak pomimo faktu, że IRSNB 3119 jest najbardziej kompletną kręgosłupem Mosasaurus lemonnieri , liczba kręgów w innych próbkach należących do tego gatunku może być znacząco różna (u okazu IRSNB 3211 zachowuje 20 kości krzyżowych, 40 pośrednich kręgów ogonowych i 31 końcowych kręgów ogonowych, u okazu IRSNB 3210 zachowuje 22 kręgi krzyżowe, 15 pośrednie ogonowe i 32 końcowe kręgi ogonowe; Lingham-Soliar, 2000). Obie próbki są niekompletne, ale zachowują większą liczbę kręgów trzonu ogonowego niż obserwuje się w przegubowej kolumnie kręgowej IRSNB 3119 (Lindgren i współpracownicy, 2011; Lingham-Soliar, 2000).
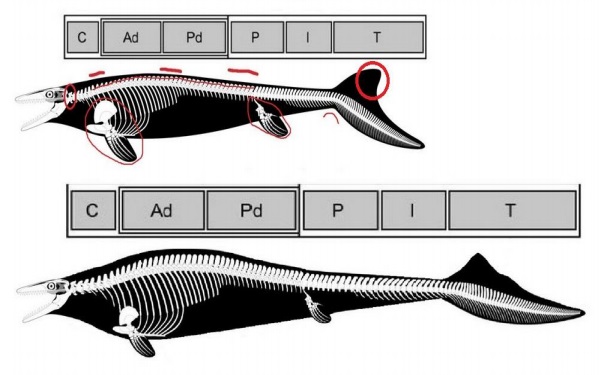
Pod koniec roku 2017 ukazała się rekonstrukcja Mosasaurus hoffmanni CCMGE 10/2469 przez pewien czas uważana za najbardziej prawidłową, której podjął się użytkownik portalu Deviantart PWNZ35-Dragon. Jednak wraz z upływającym czasem na paleoblogach osoby pasjonujące się wymarłymi gatunkami zwierząt morskich dostrzegali i wytykali tej rekonstrukcji coraz więcej błędów (niektóre z błędów zaznaczone kolorem czerwonym). Najbardziej znaczącymi z nich są: kolczaste procesy kręgów szyjnych i piersiowych są o ponad połowę niższe niż powinny ( powołując się na Lingham Soliar, 1995; Street i Caldwell, 2017) / Kręgosłup jest nieprawidłowo podzielony i zniekształcony / Odcinek kręgów szyjnych jest krótszy niż powinien / Odcinek piersiowy jest dłuższy a lędźwiowy krótszy / Długość ogona jest zbyt krótka (w tym kręgi ogonowe grupy Pygal są około dwa razy krótsze niż powinny) / Płetwa ogonowa jest zbyt mocno zakrzywiona w dół (powołując się na Lindgren, 2011) / Przednie i tylne płetwy są zbyt duże (powołując się na Lingham Soliar, 1995 i Street & Caldwell, 2017).
U Plotosaurus bennisoni liczba kręgów wynosi około 134-140 (8-10 szyjnych, około 35 grzbietowych, 28-37 krzyżowych, 3-6 pośrednich ogonowych i około 55 końcowych kręgów ogonowych; Lindgren i in współpracownicy, 2008). Najwyraźniej oczekiwana ‘’formuła kręgowa’’ Mosasaurus hoffmanni to 7 kręgów szyjnych (standardowa liczba dla przedstawicieli rodzaju Mosasaurus , chociaż nie jest wykluczona jest liczba o 2 –3 kręgi więcej jak u wcześniej wspomnianego Plotosaurus bennisoni, który jest najbliższy filogenetycznie w stosunku do Mosasaurus hoffmanni (Simões i współpracownicy,2017) 33-40 kręgosłupa, 11-13 kręgów krzyżowych, 24-25 pośrednich kręgów ogonowych (tj. około 35-38 kręgów w rdzeniu ogonowym, chociaż może być ich około 40 lub więcej w oparciu o maksymalną możliwą liczbę kręgów ogonowych u Plotosaurus bennisoni i Mosasaurus lemonnieri ), a także 49 -55 lub więcej terminalnych kręgów ogonowych. Ponadto przedstawiciele rodzaju Mosasaurus i Plotosaurus mogą różnić się proporcjami kręgów, np. Lindgren i współpracownicy 2011 wskazali, że ogon UCMP 61221 (nieokreślony okaz mozazaura z kręgami podobnymi do Mosasaurus conodon ) zajmował około 120% długości części przed kręgowej kręgosłupa, pomimo niewielkiej liczby kręgów ogonowych. Russel (1967) zauważył, że tylne kręgi ogonowe to Mosasaurus conodon są owalne w kierunku poziomym, w przeciwieństwie do owalu w kierunku pionowym tylnych kręgów ogonowych u Mosasaurus missouriensis. Ogólnie rzecz biorąc, oczekiwane rozmiary Mosasaurus hoffmanni są wysoce zależne od liczby kręgów kręgosłupa, które można przywrócić na podstawie Mosasaurus missouriensis, Mosasaurus lemonnieri lub Plotosaurus bennisoni.
Ponieważ rozmiar kręgów ogonowych może znacznie różnić się w kręgosłupie, długość części ogonowej kręgosłupa należy rozpatrywać osobno. U najbardziej bliskich krewnych Mosasaurus hoffmanni ( Plotosaurus bennisoni, Mosasaurus missouriensis i Mosasaurus sp. UCMP 61221) długość końcowych kręgów ogonowych, pośrednich ogonowych i krzyżowych wynosi ~ 120% długości odcinka kręgosłupa grzbietowego i szyjnego (Lindgren i współpracownic,2011). Z drugiej strony, ogon najbardziej kompletnego okazu Mosasaurus lemonnieri , IRSNB 3119, jest najwyraźniej proporcjonalnie nieco krótszy, zajmując około 110% długości przed kręgowej części kręgosłupa i około 45% całkowitej długości zwierzęcia, biorąc pod uwagę stosunkowo małą czaszkę (Dollo, 1892). W tym przypadku IRSNB 3119 ma porównywalną liczbę kręgów ogonowych z Plotosaurus bennisoni i Mosasaurus missouriensis (38 w porównaniu z 35 i 34-40 w łodydze ogonowej, 49 w porównaniu z 55 lub więcej w płetwie ogonowej), a niektóre inne próbki Mosasaurus lemonnieri zachowują więcej kręgów, przynajmniej w ogonie. Street & Caldwell (2016) podają około 11,4 cm długości kręgów kręgosłupa i około 11,1cm długości kręgów szyjnych dla okazu NHMM 006696, niestety położenie tych kręgów jest nieznane, a podobne dane dla innych próbek Mosasaurus hoffmanni niedostępne w literaturze. W przypadku bardzo przybliżonych oszacowań wielkości Mosasaurus hoffmanni możemy jedynie założyć, że kręgi te miały „średnią” długość w pełnej kolumnie kręgosłupa.
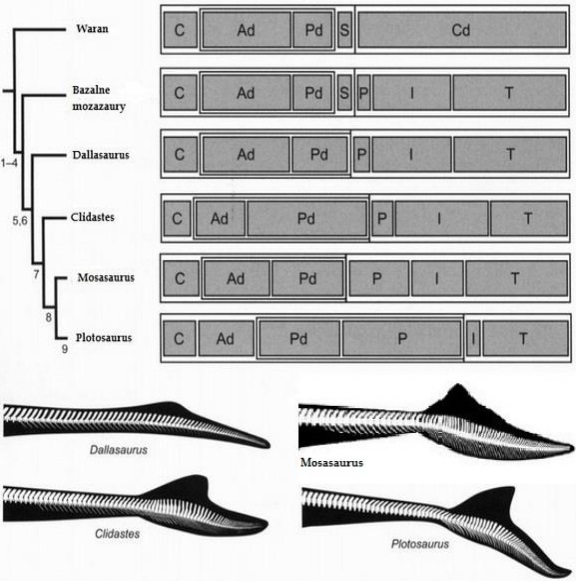
Proporcje poszczególnych odcinków kręgosłupowych u różnych przedstawicieli mozazaurów oraz ich rekonstrukcje ogonowe. C – kręgi szyjne: Ad – kręgi piersiowe: Pd – kręgi lędźwiowe: S – kość krzyżowa: Cd – kręgi ogonowe: P -kręgi z grupy Phygal: I -kręgi pośrednie ogonowe: T- końcowe kręgi ogonowe.
Zatem odcinek kręgu szyjnego NHMM 006696 będzie miała około 77,7 cm długości. Część grzbietowa kręgosłupa, w oparciu o liczbę kręgów grzbietowych TMP 2012.010.0001 (Mosasaurus missouriensis ), będzie miała długość około 376,2 cm, wskazując całkowitą długość części przed kręgowej kręgosłupa na około 453,9 cm i pełnej długości kręgosłupa w okolicy 998,5 cm (długość ogona = 120% długości tułowia i szyi). Część grzbietowa kręgosłupa w oparciu o liczbę kręgów grzbietowych Plotosaurus bennisoni będzie miała długość około 433,2 cm, co wskazuje całkowitą długość części przed kręgowej kręgosłupa około 510,9 cm i długość pełnej kręgosłupa w okolicy 1124 cm (długość ogona = 120% długość ciała i szyi). Część grzbietowa kręgosłupa, w oparciu o liczbę kręgów grzbietowych IRSNB 3119 (Mosasaurus lemonnieri ), będzie miała długość około 456 cm, co wskazuje całkowitą długość części przed kręgowej kręgosłupa w przybliżeniu na ok. 533,7 cm i długość pełnej kręgosłupa w okolicy 1120,8 cm (długość ogona = 110% długości tułowia i szyi). W przypadku czaszki z żuchwą od długości 144 cm rekonstrukcje powinny wskazywać całkowitą długość NHMM 006696 w obszarze odpowiednio 1133,5 cm, 1259 cm i 1255,8 cm, stosunek długości dolnej szczęki do całkowitej długości w zakresie 1: 7,87, 1: 8,74 i 1: 8,72 (wartość średnia ~ 1: 8,6).
Skalowanie okazu CCMGE 10/2469 (największa ze stosunkowo kompletnych znanych obecnie szczęk dolnych o długości 171 cm), daje całkowitą długość 1345,8 cm, 1494,5 cm i 1491 cm (wartość średnia = 1443,8 cm). Liczby te zakładają całkowitą długość wzdłuż krzywej i nie uwzględniają długości przestrzeni międzykręgowej. Wczesne szacunki sugerowały, że długość krążków międzykręgowych u dużych przedstawicieli mozazaurów (np. Tylosaurus proriger KUVP 5033) wynosiła do 1,3 cm lub więcej (Everhart, korespondencja osobista). Jeśli Mosasaurus hoffmanni miał taką samą długość krążków międzykręgowych (należy zauważyć, że kręgi największych tylozaurów, takie jak KUVP 5033, są porównywalne pod względem wielkości do kręgów NHMM 006696), całkowitą długość NHMM 006696 na podstawie TMP 2012.010.0001 (Mosasaurus missouriensis ), Plotosaurus bennisoni i IRSNB 3119 ( Mosasaurus lemonnieri ) można zwiększyć o 13- 14%. Jednak obecnie uważa się, że mozazaury nie miały krążków międzykręgowych, a ich kręgi były połączone raczej jak stawy, jak w przypadku współczesnych waranowatych, zachowując jedynie niewielką przestrzeń dla płynu międzykostnego niezbędnego do ruchu kręgów (Rothschild i Everhart, korespondencja osobista). W tym przypadku długość przestrzeni między kręgami wynosiła ok 0,57 cm dla dużych okazów, takich jak NHMM 006696
(Everhart, Street, Schulp, Lindgren, korespondencja osobista). Korzystając z proporcji najbliższych filogenetycznych krewnych Mosasaurus hoffmanni, Plotosaurus bennisoni, Mosasaurus missouriensis i Mosasaurus lemonnieri i bardzo przybliżonych obliczeń kręgów można dojść do wniosku, że najbardziej prawdopodobny stosunek długości dolnej szczęki do całkowitej długości Mosasaurus hoffmanni wśród możliwych opcji (1: 7,2, 1: 8,14, 1: 8,5, 1: 9,02, 1: 9,03, 1; 9,5 i 1:11) wynosi 1: 8,91. Zatem całkowita długość CCMGE 10/2469 (żuchwa długości 171 cm) wynosiła prawdopodobnie około 15,24 metra.Chociaż nie zaproponowano żadnych poważnych szacunków masy ciała Mosasaurus hoffmanni najwyraźniej nie odpowiadał historycznym wyobrażeniom o mozazaurach jako serpentynowatych zwierzętach morskich i najprawdopodobniej charakteryzowały się dość gęstą budową ciała. Klatka piersiowa Mosasaurus hoffmanni ma kształt beczki, jest głęboka i stosunkowo szeroka w porównaniu do na przykład innego przedstawiciela mozazaurów Tylozaura (Lingham-Soliar,1995), który miał klatkę piersiową ściśniętą bocznie. Żebra były grubsze niż u Hainosaurus czy Plotosaurus. Płetwy były stosunkowo krótkie za to dość szerokie. Ogromny tułów przypominający ichtiozaury, delfiny i rekiny jest również sugerowany dla Plotosaurus bennisoni (Lindgren i współpracownicy., 2007) i jest również obserwowany w dobrze zachowanej próbce Mosasaurus missouriensis (Konishi i współpracownicy, 2014). Zakładając, że gęstość ciała Mosasaurus hoffmanni odpowiada typowej gęstości dla zwierząt morskich wynoszących 1kg/l, w oparciu o spodziewaną objętość ciała, masę 1524 cm Mosasaurus hoffmanni można oszacować na około 18869 kilogramów.
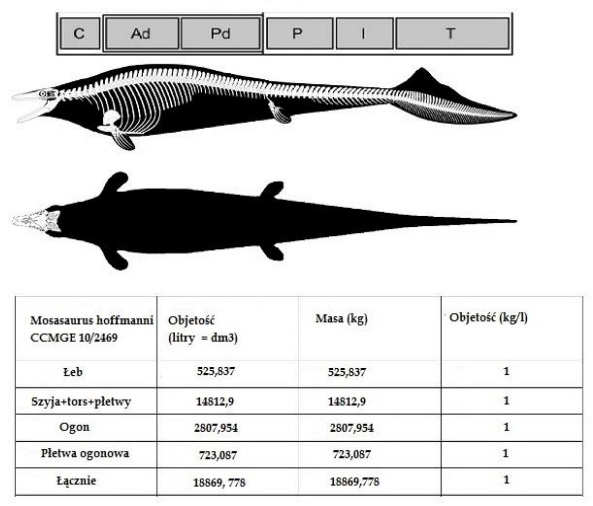
Czaszka na podstawie rekonstrukcji z Street i Caldwell (2017). Liczba kręgów na podstawie prawie kompletnego szkieletu Mosasaurus missouriensis (TMP 2012.010.0001), co daje dość zachowawcze oszacowanie (dla CCMGE 10/2469 z 171 cm żuchwą) 1524 cm długości. Kręgi szyjne i grzbietowe na podstawie MNHN AC 9648, IRSNB R26 i NHMM 006696 z dokumentów Lingham-Soliar (1995) i Street & Caldwell (2017), a także na TMP 2012.010.0001. Ogon całkowicie oparty na TMP 2012.010.0001. Pas piersiowy, biodrowy, przedni i tylny na podstawie IRSNB R26, MNHN AC 9648, NHMM 1193024 i NHMM 006696. Klatka piersiowa dopasowana od TMP 2012.010.0001. Elementy mostkowe spekulatywne. Kontrowersyjną częścią odbudowy jest wielkość kończyn. Użyłem danych od Anne Schulp, a nie skalowania kości kończyn IRSNB R12 (Lingham-Soliar,1995), ponieważ ten ostatni okaz wydaje się być młodym osobnikiem, który może mieć nieproporcjonalnie krótkie kończyny. Płetwa ogonowa oparta jest na rekonstrukcji Lindgren i współpracownicy,2011).
W tym miejscu praktycznie kończąc artykuł chciałbym dodać jeszcze informację na temat odkryć przypisywanych do Mosasaurus hoffmanni na przestrzeni lat i historii z nią związanych oraz dane szacunkowe dotyczące szacunkowej długości ciała i masy poszczególnych skatalogowanych gatunków Mosasaurus hoffmanni.
W latach 1770 –1774 emerytowany lekarz wojskowy Johann Leonard Hoffman dokonał odkrycia drugiej fragmatycznej czaszki na ziemi należącej do Theodora Junana Goddinga, którą później przekazał holenderskiemu profesorowi Peterowi Kamperowi. Początkowo Hoffman zakładał, że znaleziona przez niego czaszka należy do krokodyla, ale Camper w 1786 roku doszedł do wniosku,
że należała ona do nieznanego rodzaju kaszalota.W 1794 roku miasto Maastricht zostało przejęte przez francuską armię rewolucyjną, dwa miesiące później przybył tam geolog Barthélemy Faujas de Saint-Fond w towarzystwie żołnierzy, gdzie
skonfiskował znalezisko uznając ją za bardzo wartościową naukowo i w grudniu 1794 roku przetransportował ją do Paryża jako trofeum wojenne i dodana do kolekcji Narodowego Muzeum
Historii Naturalnej. De Saint-Fond również uważał, że próbka była krokodylem, ale w 1798 roku syn Petera Kampera, Adrian ponownie zbadał skamielinę dokonując korekty opisu swojego ojca, będąc pierwszym, który doszedł do wniosku, iż skamieniałość należy do wymarłego zwierzęcia będącego spokrewnionym z jaszczurkami z rodziny waranowatych (później tę opinię potwierdził G. Cuvier, który korespondował z Camperem Jr.).W 1927 roku odkryta została fragmatyczna czaszka Mosasaurus w pobliżu miasta Penza (Rosja), która była pierwszym udokumentowanym zapisem odkrycia Mosasaurus z porewolucyjnej Rosji.
Cześć pierwotnie znalezionych kości została następnie utracona, a ich opis został dokonany na odlewach gipsowych. Kopia Penza jest jednym z największych Mozazaurów jakie kiedykolwiek
odnaleziono i jest wyjątkowa zarówno pod względem i okoliczności odkrycia. W 1927 roku wygnaniec polityczny MA Vedenyapin znalazł kość dużego gada morskiego na obrzeżach Penza w wąwozie, w którym trenowano żołnierzy Armii Czerwonej. Cała populacja zaczęła mówić o wykopywaniu przedpotopowego zwierzęcia. Podczas kolejnych wykopalisk znaleziono kości żuchwy, łopatki, kręgi i żebra. Aby zapewnić zachowanie znalezionego materiału, umieszczono je w skrzynkach i wraz z blokami ziemi, które następnie wysłano do Komitetu Geologicznego St. Petersburga. Oryginalna czaszka okazu z Penzy jest częścią eksponatów w hali wystawowej Centralnego Naukowo-Badawczego Muzeum Geologiczno-Eksploracyjnego imienia F.N. Czernyszewa w Petesburgu, kopia jest natomiast jest częścią wystawy muzeum w Penzie. Niestety, czaszka wystawiona w Petersburgu ‘’cierpiała’’ na taki sam los, jak niektóre kości podczas wykopalisk mianowicie wszystkie małe, luźne kości i zęby zostały skradzione przez odwiedzających muzeum, więc w późniejszym czasie dyrekcja podjęła decyzje o umieszczeniu czaszki za szklana kopułą i zastąpieniu skradzionych oryginalnych zębów kopiami.10 września 2012 roku w kamieniołomie wapienia pod Maastricht, ponownie znaleziono kości Mosasaurus. Był to ten sam kamieniołom, w którym znaleziono typową próbkę Mosasaurus
hoffmanni. Dziesięć dni później pracownicy muzeum odkryli jeszcze kilka dużych fragmentów czaszki, kości ciała i ogona, szacując długość zwierzęcia na nieco ponad 12 metrów. Wiek okazu
oszacowano na 67,83 milionów lat, co czyni go o około półtora miliona lat starszym od innego wielkiego przedstawiciela mozazaurów Prognathodon o ksywie ‘’Ber’’. Wydaje się, że jest to jeden z najstarszych znanych okazów Mosasaurus hoffmanni , chociaż szczegółowe badania nie zostały jeszcze przeprowadzone i możliwe, że jest to inny gatunek lub nawet przedstawiciel innego rodzaju.W 2014 r. Interesujące odkrycie zostało opisane przez Takuyę Konishi i współpracowników w czasopiśmie Vertebrate Paleontology. Historia ta rozpoczęła się w 2008 roku, gdy pracownicy
kanadyjskiej firmy Korite International Ammolite Mine, zajmującej się wydobyciem rudy amonitu, odkryli kości gadów morskich. Paleontolodzy z Muzeum Paleontologii Royal Tyrrell odzyskali
szkielet, ale dopiero po starannym przygotowaniu i badaniu stało się jasne, że odkryli cenną skamielinę. Ten Mosasaurus został zachowany wraz z jego ostatnim posiłkiem w żołądku, który stanowiła ryba kostna o wielkości ok. Metra oraz ‘’pozwolił’’ autorom badania zidentyfikować chrząstkowe pierścienie tchawicy, a także mostka. Chociaż brakuje niektórych części tylnej ciała, reszta szkieletu jest bardzo dobrze zachowana. Na jego kościach znaleziono również ślady ugryzień co najmniej trzech różnych rekinów.W kwietniu 2015 roku paleontolog amator Lars Barten z Ridgekevurt w Holandii wraz z ojcem Jos Bartenem odkryli skamieniałości Mosasaurus hoffmanni , okaz został nazwany ‘’ Lars’’ od imienia odkrywcy i znajduje się obecnie w Muzeum Historii Naturalnej w Maastricht.
*Opublikowany artykuł może być wykorzystywany wyłącznie na własny użytek edukacyjny, nie może stanowić źródła informacyjnego dla innych blogów, wykluczonym jest także udostępnianie ani publikowanie go bez zgody autora.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Poprzedni artykuł:
Walka o tron Następny artykuł:
Oszacowania jak duże były kenozoiczne krokodyle i krokodylomorfy7 odpowiedzi do: "Mosasaurus hoffmanni"
Dodaj komentarz
-






































Jak zwykle, artykuł wypas, dzięki Krzysiek!
Krzysztof, fajny artykuł, jak zwykle. Mam jedną, drobną sugestię, może będzie pomocna. Moim zdaniem fajnie byłoby zamieścić, taki akapit podsumowujący wielkość opisywanego gada. Przeczytałem z uwagą cały artykuł i o ile widzę w tabeli szacunki masy osobników od 815 kg do 18.8 tony, to nie do końca wiem, ile mógł mierzyć największy odkryty osobnik – 15.24 metra?. Dodałbym coś takiego, jak przy opisach stosowane jest w artykułach w Dinoanimals, długość i zakres, masa i zakres, oczywiście z uwzględnieniem całości artykułu. Jeden rzut oka wystarczyłby aby to ogarnąć 🙂
Duży był ale przy dzisiejszych płetwalach błękitnych to maluch 😉
A mozazaury żyły tylko w kredzie czy już wcześniej?
Mozazaury żyły tylko w kredzie.
Perfekcyjny artykuł jak zawsze Krzychu (już myślałem,że zrezygnowałeś z tworzenia artykułów na DA), jakieś plany na kolejne artykuły na blogu ?. Tylko oby przerwa nie trwała 5 miesięcy tak jak w przypadku pomiędzy poprzednimi artykułami.
Ciekawy wpis