
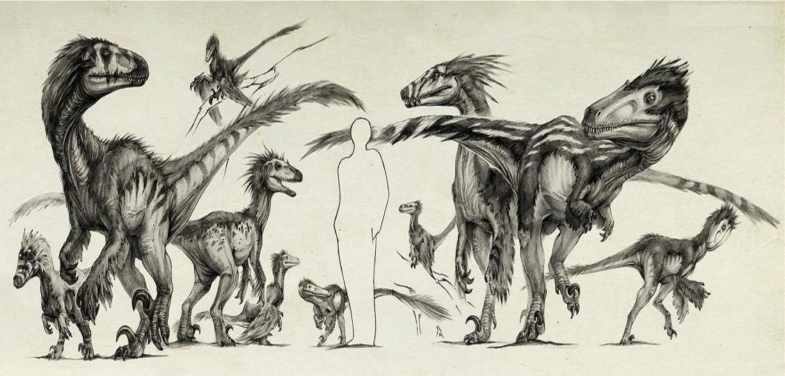
W swoim dzisiejszym artykule chciałbym wam przedstawić piątkę największych przedstawiciel dromaeosauridae ( które personalnie są moją ulubioną rodziną teropodów ).

Nazwa: Utahraptor ostrommaysorum (rabuś/łowca z Utah – epitet honoruje paleontologa Johna Ostorma – odkrywcy Deinonychusa i Chrisa Maysa, szefa Dinamation International Corporation, który zorganizował badania w Dalton Wells). Nazwa nadana przez Jamesa Kirklanda, Roberta Gastona oraz Donalda Burgea w 1993 roku.
Specyfikacja: Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce- Podtyp: kręgowce Gromada: zauropsydy – Podgromada: diapsydy – Infragromada: archozauromorfy – Nadrząd: dinozaury – Rząd: dinozaury gadziomiednicze – Podrząd: teropody – Infrarząd: tetanury (bez rangi: celurozaury – Nadrodzina: deinonychozaury – Rodzina: dromeozaury – Rodzaj: Utahraptor – Gatunek: Utahraptor ostrommaysorum.
Szacunek masy i długości: W pełni dorosły osobnik Utahraptora osiągał 7 metrów długości (w tym 71 cm przypadało na czaszkę ), masę ok. 500 kilogramów ( niektórzy paleontolodzy sądzą, że masa dorosłych osobników mogła dochodzić nawet do 700 kilogramów ) oraz ok. 2,2 metra wysokości. Jako ciekawostkę można podać, że odnaleziono pojedynczy kręg ogonowy Utahraptora. Na jego podstawie przypisuje się właścicielowi ogromne rozmiary 11 – 12 m długości i 2,7 t masy! Należy pamiętać przy tym jednak, że owa skamielina nie została formalnie opisana a obliczenia rozmiarów na podstawie pojedynczej kości są mocno niepewne i często błędne.
Czas i miejsce występowania: Stany Zjednoczone (Utah), formacja Cedar Mountain – ogniwa Yellow Cat i Ruby Ranch – ok. 124- 113 mln lat temu wczesna kreda (apt).
Odkrycie: Pierwsze okazy Utahraptora zostały znalezione w 1975 roku przez Jima Jensena w Dalton Wells Quarry w środkowo-wschodnim Utah, niedaleko miasta Moab.
Klasyfikacja: Budowa pazurów i kręgów silnie wskazuje na przynależność Utahraptora do rodziny Dromaeosauridae. Serracja na zębach bardziej przypomina tę, która występuje u Dromaeosaurinae niż Velociraptorinae. Potwierdza to m. in. praca Turnera i in. (2007), wg której Utahraptor jest blisko spokrewniony z Achillobatorem,Dromaeosaurusem oraz Adasaurusem.
Reprezentacja skamieniałości: Liczne okazy szczątków fragmentarycznych.
Charakterystyka: Utahraptora, można uznać za wcześniejszą powiększoną formę Deinonychusa , i choć szczątki tego dinozaura są fragmatyczne to naukowcy szacują , że dorosłe osobniki osiągały 7 metrów długości i 500 kg co sprawia, iż Utharaptor uznawany jest za największego przedstawiciela dromeozaurów. Utharaptor jako największy okaz wcześniej wspomnianych posiadał największy zakrzywiony ”sierpowaty pazur” o długości 24 centymetrów (wartość mierzona po krzywej), jego potężniej zbudowane kości i włókna mięśniowe kończyn dolnych wskazywały na to, że był szybkim myśliwym w związku z tym powyższym Utharaptor , był on w stanie bez problemu dogonić większe ofiary w postaci hadrozaurów czy mniejszych zauropodów. Jako przedstawiciel dromeozaurów Utahraptor prawdopodobnie wykazywał wszystkie cechy filogenetyczne grupy łącznie z domniemanym upierzeniem. Warto tu jednak wspomnieć, iż duże gabaryty tego teropoda zdaniem niektórych paleontologów wskazują na jego potencjalną zmiennocieplność, która nie występowała jako cecha u jego mniejszych krewnych (jeżeli ta teoria była by trafna to Utahraptor charakteryzował by się mniejszym stopniem upierzenia lub innym jego rozkładem).

Nazwa: Austroraptor cabazai (nazwa oznacza południowego rabusia nazwa odnosi się do miejsca odkrycia Ameryki Południowej – epitet gatunkowy nadano dla uczczenia pamięci zmarłego Héctora Cabazy, założyciela Museo Municipal de Lamarque). Nazwa nadana została przez Novasa, Pola, Canalea, Porfiria i Calvo w 2009 roku.
Specyfikacja: Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce – Gromada: zauropsydy – Podgromada: diapsydy – Infragromada: archozauromorfy – Nadrząd: dinozaury – Rząd: dinozaury gadziomiednicze – Podrząd: teropody – Infrarząd: tetanury (bez rangi: celurozaury) – Nadrodzina: deinonychozaury – Rodzina: dromeozaury – Podrodzina: unenlagie – Rodzaj: Austroraptor – Gatunek: Austroraptor cabazai.
Szacunek masy i długości: Dorosły osobnik Austroraptora określany jest przeważnie na 5 metrów długości (z czego na czaszkę przypada 80 cm), Novasa i współpracownicy (2009) kwestionują jednak ów wartość uznając ją za zaniżoną, takiego samego zdania jest Vega (online 2008), którego skrupulatne obliczenia przypisują Austroraptorowi dokładną długość 6,8 metra, natomiast Gregory S. Paul (2010) szacuje jego długość na 6 metrów i ciężar ciała o wartości minimalnej nie mniejszej niż 300 kg. Masa ciała przeważnie określana jest w przedziale od 370 do 520 kilogramów oraz wysokość ok. 1,6 – 1,8 metra.
Czas i miejsce występowania: Argentyna – prowincja Rio Negro, formacja Allen ok. 70 mln lat temu późna kreda (mastrycht).
Odkrycie: Holotyp MML-195, został odnaleziony w miejscowości Bajo de Santa Rosa formacji Allen, w Río Negro w Argentynie w 2002 roku przez zespół prowadzony przez Fernando Emilio Novasa z Museo Argentino de Ciencias Naturales.
Klasyfikacja: Analiza cech anatomicznych umieszcza Austroraptora w podrodzinie Unenlagiinae (Turner, Makovicky i Norellin 2012). Austroraptor był blisko spokrewniony z Buitreraptorem.
Reprezentacja skamieniałości: Holotyp MML-195 w skład, którego wchodzą: kości czaszki: czołową, zaoczodołową, kości łzowe, szczękowe i zębowe z zębami, kość kątową górną oraz część kości zębowej, poprzedzającej kość stawową. Na materiał pozaczaszkowy składają się: kręgi szyjne (3, 5, 6, 7 i 8), kręgi grzbietowe (2 i 4), izolowane żebra, także brzuszne, prawa kość ramienna, pazur (III) z kończyny górnej, lewy trzonek łonowy oraz kości kończyn tylnych: lewa kość udowa, prawa kość piszczelowa, kość skokowa, piętowa, kość śródstopia (III) oraz paliczki I-2, II-2, III-4 i IV-2).
Drugi okaz (MML-220) to niekompletna (n.) kość szczękowa, dwa zęby, n. kręg grzbietowy, 12 tylnych kręgów ogonowych, n. k. ramienna, kk. promieniowe, łokciowe (n.), k. śródręcza I, paliczek I-1, n. pazur dłoni I, oba pazury II, n. pazur III, piszczel, paliczek stopy I-1, n II k. śródstopia, paliczek II-1, n. III k. śródstopia, paliczek III-1, 2 i 3 paliczek IV-1, 2, pazur IV.
Charakterystyka: Choć nie tak duży jak Północno Amerykański Utahraptor , Austroraptor nadal pozostaje w ścisłej czołówce największych przedstawicieli dromeozaurów ( największy obecnie znany dromeozauroid Ameryki Południowej). Teropod ten wykazuje wiele funkcji, które czynią go unikalnym na tle innych członków grupy, pierwsza są proporcjonalnie małe przedramiona, które zazwyczaj są dobrze rozwinięte u innych dromeozaurów dzięki czemu mogły one w skuteczniejszy sposób przytrzymywać zdobycz by następnie zadawać jej rany przy użyciu pazurów kończyn dolnych. Drugim czynnikiem jest dość wąska i wydłużona czaszka co wskazuje na brak dostatecznie rozwiniętych ” mięśni gryzących ” co naturalnie oznacza proporcjonalnie mniejszą siłę ugryzienia. Wreszcie zęby, które nie są tak silne i nie wykazują tak silnej seracji jak u innych dromeozaurów a są bardziej stożkowane nadające się bardziej do przytrzymywania ofiary co może sugerować to, iż Austroraptor mógł być niszowym drapieżnikiem skupiającym się na jednym typie łupu.
Niedostatecznie rozwinięte ramiona oznaczały, dinozaur ten nie był w stanie przytrzymywać większej ofiary i polował na mniejszą zwierzynę, także stożkowane zęby budowa anatomiczna przypominające te jakie posiadały spinozaury (grupa wyspecjalizowanych teropodów uważanych przede wszystkim za rybożerców) oraz słabszy nacisk szczęki sugerowały by właśnie w przypadku Austroraptora specjalizacje w polowaniu na mniejsze ofiary lub nawet rybożerności, gdzie tak jak spinozaury używał by on swoich pazurów do rozszarpywania ryb. Nie wyklucza się też jego potencjalnej padlinożerności, gdzie wykorzystując swój rozmiar odpędzał by mniejsze drapieżniki od upolowanego przez nie uprzednio łupu.
Mimo, iż sierpowate pazury dromeozaurów są bardziej kojarzone z funkcją kłucia/dźgania/ niż z cięciem/przebijaniem/przecinaniem to nadal mogły one skutecznie wyrywać kawałki ciała mniejszych zwierząt tak aby słabsze szczęki i stożkowate zęby i słabsze mięśnie szczek nie stanowiły problemu w efektywnym odżywianiu. Jeśli wcześniejsza koncepcja była by faktycznie poprawna to technika łowiecka Austroraptora opierała by się bardziej na szybkości/zwinności i wytrzymałości niż siły. Austroraptor mógł również używać pazurów do uderzania w ryby znajdujące się w płytkich wodach, gdzie mógł je następnie wyłowić wyspecjalizowanymi zębami trzymając je w pysku, a na brzegu rozszarpywać je na mniejsze kawałki.

Nazwa: Achillobator giganticus (nazwa nawiązuje do postaci Achillesa herosa znanego z „Iliady” Homera, epitet gatunkowy tyczy się rozmiaru zwierzęcia. Samo słowo bator w języku mongolskim oznacza bohatera). Nazwa została nadana przez Altangerela Perlea, Marka A. Norella i Jima Clarka w 1999 roku.
Specyfikacja: Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce – Gromada: zauropsydy – Podgromada: diapsydy – Nadrząd: dinozaury – Rząd: dinozaury gadziomiednicze – Podrząd: teropody – Infrarząd: tetanury (bez rangi: celurozaury) – Nadrodzina: deinonychozaury – Rodzina: dromeozaury – Rodzaj: Achillobator – Gatunek: Achillobator giganticus.
Szacunek masy i długości: Dorosły osobnik Achillobatora osiągał 5 – 6 metrów długości, masa ciała szacowana jest minimalnie na 80 kilogramów a maksymalnie na 250 kilogramów (287 – 320 kg ?) oraz ok.1,6 -1,7 metra wysokości.
Czas i miejsce występowania: Mongolia prowincja Dornogovi, formacja Bayan Shireh ok.98 – 83 mln lat temu późna kreda (cenoman – santon).
Odkrycie: Skamieniałości Achillobatora po raz pierwszy przez Mongolsko/Rosyjską ekspedycję w obszarze Burkhant w 1989 roku.
Klasyfikacja: Analizy filogenetyczne przeprowadzone przez Longricha i Currie (2009) oraz Senter i współpracowników (2012) pokazują, że Achillobator jest członkiem podrodziny Dromaeosaurinae, najbardziej spokrewnionym z formami Ameryki Północnej, takimi jak Utahraptor, Deinonychus, Velociraptor oraz Dromaeosaurus .
Reprezentacja skamieniałości: Holotyp (FR. MNUFR-15) (ok. 5 m dł., osobnik dorosły): kość szczękowa (290,8 mm), 9 zębów (do 38 mm), szósty kręg szyjny (34,6 mm), dziesiąty kręg szyjny (51,3 mm), czwarty (?) kręg grzbietowy (53,8 mm), tylny kręg ogonowy (49,5 mm), przednie i tylne żebro, środkowy krąg ogonowy, 6 dystalnych kręgów ogonowych, 3 szewrony, kości łopatkowa i krucza, kość promieniowa (260 mm), kość śródręcza III (71 mm), paliczek I-1 (78 mm), 2 pazury dłoni (112 mm), kość biodrowa (531 mm), kości łonowe (548 mm), kulszowa (378 mm), udowa (505 mm), piszczelowa (490,4 mm), paliczki II-2 (56,4 mm), III-2 (55 mm), kości śródstopia III (234,4 mm) i IV (209,6 mm).
Charakterystyka: Choć odkryty już w 1989 roku Achillobator nie posiadał oficjalnej nazwy aż do 1999 roku i choć jego szczątki są fragmatyczne to jednak w kręgu naukowym nie ma wątpliwości co do tego, że był on jednym z największych przedstawicieli dromeozaurów. Ścięgna Achillesa tego teropoda była wyjątkowo dobrze rozwinięte co w pewnym kontekście stanowiło podwaliny do powołania nazwy rodzaju. Achillobator w przeszłości ”został oskarżony” hipotezę chimery co w najprostszych słowach wyjaśnienia oznacza, że materiał kopalny przypisany do rodzaju faktycznie reprezentuje więcej niż jeden rodzaj dinozaura, głównym poparciem tej teorii jest to, że kość łonowa (najbardziej wysunięta do przodu biodra) wskazywała pozycję pionową w dół, podczas gdy u wszystkich innych przedstawicieli kość ta skierowana jest ku tyłowi i prowadzi do łuku wskazującego w tym samym kierunku co kość kulszowa (kości w tylnej części bioder). Opinia ogólna wskazuje dziś Achillobatora jako dromaeosaurida z unikalną strukturą budowy biodra.
Inne cechy autapomorficzne wyróżniające Achillobatora:
• łuk kości przedszczękowej całkowicie odsłonięty
• kość przedszczękowa jest wydłużona pionowo ukierunkowane na tym samym poziomie w szczęce
• kości śródstopia są ułożone proksymalnie
• kość udowa jest dłuższa niż piszczel
• mięsień półbłoniasty jest duży i trójkątny w budowie i umiejscowiony w połowie kości kulszowej

Nazwa: Dakotaraptor steini (nazwa oznacza rabusia z Dakoty i odnosi się do miejsca znalezienia szczątków omawianego teropoda, epitet gatunkowy honoruje paleontologa Waltera Steina). Nazwa została nadana przez Roberta A. DePalme, Davida A. Burnhama, Larryego Dean Martina, Petera Lars Larsona& Roberta Thomasa Bakkera w 2015 roku.
Specyfikacja: Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce – Gromada: zauropsydy – Podgromada: diapsydy – Infragromada: archozauromorfy- Nadrząd: dinozaury – Rząd: dinozaury gadziomiednicze – Podrząd: teropody – Infrarząd: tetanury (bez rangi: celurozaury) – Nadrodzina: deinonychozaury – Rodzina: dromeozaury – Rodzaj: Dakotaraptor – Gatunek: Dakotaraptor steini.
Szacunek masy i długości: Dorosły osobnik Dakotaraptora szacowany jest na 5,5 metra długości, masę ciała w przedziale 220 – 250 kilogramów (453 kg -?) i 2 metry wysokości.
Czas i miejsce występowania: Stany Zjednoczone Dakota Południowa – Harding, formacja Hell Creek ok. 67 – 66 mln lat temu późna kreda (późny mastrycht).
Odkrycie: Szczątki holotypu Dakotaraptora zostały znalezione w piaskowcach górnych warstw formacji Hell Creek w stanie Dakota Południowa w hrabstwie Harding, odkrycia dokonał paleontolog Robert DePalma w 2005 roku.
Klasyfikacja: Pomimo, że Dakotaraptor był jednym z największych przedstawicieli Dromaeosauridae, to jednak proporcje jego ciała bardziej przypominały te występujące u mniejszych krewniaków, jak Dromaeosaurus czy Deinonychus. Analiza filogenetyczna wykonana przez autorów opisu sugeruje, że najbliższym krewnym Dakotaraptora był Dromaeosaurus albertensis. Inny wynik dała niepublikowana analiza Cau (2015 online). Wg tego włoskiego paleontologa Dakotaraptor był taksonem siostrzanym do kładu obejmującego Dromaeosaurinae i Velociraptorinae. Z taką interpretacją zgadza się także Mortimer (2016 online).
Reprezentacja skamieniałości: Holotyp oznaczony jako PBMNH.P10.113.T obejmuje drugi i trzeci pazur prawej stopy, prawą kość udową, prawą i lewą kość piszczelową, lewą kość stępu i piętową, kości prawego i lewego śródstopia, 10 kręgów ogonowych, lewą i prawą kość ramienną, kości promieniowe, kości łokciowe, kości prawego śródręcza, fragmenty paliczków. Materiał przypisany to m.in. prawa kość piszczelowa i zęby.
Charakterystyka: Dakotaraptor pod względem anatomiczny nie różnił się od innych przedstawicieli dromeozaurów, tak jak wszystkie tak i on posiadał charakterystyczny, zakrzywiony pazur na drugim palcu stopy (długość mierzona wzdłuż krzywej grzbietowej wynosi 24 cm), pazur jest więc proporcjonalnie większy niż u Deinonychusa i przypomina pazur Utahraptora.
Kości kończyn tylnych Dakotaraptora są smuklejsze niż u będącego podobnej wielkości Utahraptora, i bardziej przypominają kości mniejszych dromeozaurydów, jak Velociraptor czy Bambiraptor. Dakotaraptor ma też proporcjonalnie dłuższe śródstopie w stosunku do łącznej długości kości udowej i piszczelowej niż Utahraptor i Deinonychus, lecz nieco krótsze niż Bambiraptora. Takie proporcje tylnych kończyn wskazują, że omawiany tu teropod był szybszym biegaczem niż np. Utahraptor.
Kości ramienna i łokciowa przypominają pod względem budowy analogiczne kości Deinonychusa. Na kości łokciowej znaleziono struktury przypominające wzgórki, na których są osadzone lotki współczesnych ptaków. Jeżeli więc wziąć by pod uwagę trafność tej hipotezy Dakotaraptor byłby największym dromeozaurydem, u którego znaleziono ślady występowania piór. Zęby są silnie spłaszczone bocznie i umiarkowanie silnie zakrzywione budową anatomiczną zbliżone są do zębów innych dromeozaurydów.

Nazwa: Deinonychus antirrhopus (nazwa Deinonychus oznacza straszny szpon, epitet gatunkowy antirrhopus natomiast przeciwwagę i odnosi się do niezwykłej w czasie nadania nazwy struktury ogona zwierzęcia). Nazwa nadana została przez Johna Ostroma w 1969 roku.
Specyfikacja: Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce – Gromada: zauropsydy – Podgromada: diapsydy – Infragromada: archozauromorfy – Nadrząd: dinozaury – Rząd: dinozaury gadziomiednicze – Podrząd: teropody – Infrarząd: tetanury (bez rangi: celurozaury) – Nadrodzina: deinonychozaury – Rodzina: dromeozaury – Rodzaj: Deinonychus antirrhopus.
Szacunek masy i długości: Dorosły osobnik Deinonychusa szacowany jest na 3,4 metra (nie wykluczone jest, że mógł osiągać większe rozmiary i masę niż wartość podana wcześniej) długości, masę w przedziale 70 – 80 kilogramów ( 100 ? ) oraz ok. 1,2 – 1,3 metra wysokości.
Czas i miejsce występowania: Stany Zjednoczone formacja Antlers – Oklahoma, formacja Cloverly – Montana i Wyoming, formacja Arundel – Maryland oraz formacja Trinity – Teksas ok. 115 – 112 mln lat temu wczesna kreda (późny apt).
Odkrycie: Pierwsze skamieniałości Deinonychusa zostały znalezione w 1931 w południowej Montanie, koło miasta Bridger przez Barnuma Browna.
Klasyfikacja: Deinonychus należy do nadrodziny Deinonychosauria, która swoją nazwę bierze od jego nazwy. Obecnie najczęściej zaliczany jest do podrodziny Velociraptorinae, która definiowana jest jako dinozaury, którym bliżej do Velociraptora niż do innych dromeozaurów. Początkowo do niej został zaliczony tylko Velociraptor, później dodano do niej również rodzaje Deinonychus i Sauronitholestes. Najnowsze badania zaliczają do tej podrodziny oprócz Velociraptora, Deinonychusa , Tsaagana i czasami Saurornitolesta.
Reprezentacja skamieniałości: Holotyp (YPM 5205) to kompletna lewa i niekompletna prawa stopa. Poza tym znanych jest wiele mniej lub bardziej kompletnych okazów, dających dobry obraz osteologii zwierzęcia, a także jaj, które – z uwagi na powiązanie ze szczątkami Deinonychów – prawdopodobnie zostały złożone właśnie przez nie.
Charakterystyka: Dinozaury przez większość osób postrzegane są jako ” wielkie ociężałe jaszczury ” przypadek Deinonychusa udowadnia całkowitą sprzeczność z ów tezą. Choć mniejszy od swoich większych pobratymców (za to szybki i stosunkowo solidnie zbudowany anatomicznie) stanowił nie mniej śmiertelne zagrożenie dla swoich potencjalnych ofiar.
Intensywniejszy tryb życia wymagał od tego drapieżnika szybszego metabolizmu, co z kolei mogło nawiązywać do jego ciepłokrwistości, co z kolei wymaga adaptacji do oszczędzania ciepła w ciele. Chociaż nie ma jednoznacznych dowodów na poparcie sugerujące by ten niewielki teropod posiadał pióra to jednak nie ma wątpliwości co do tego, że był on członkiem dromeozaurów (gdzie istnieją niezbite dowody w postaci materiałów kopalnych na obecność upierzenia), było by dość zaskakującym, gdyby Deinonychus ich nie posiadał.
Kiedy po raz pierwszy odnaleziono czaszkę Deinonychusa była ona ” rozproszona ” i w ” słabym stanie ”, gdzie ogólny początkowy jej ” obraz ” dawał wygląd nie różniący się od tej jaka posiadał Allosaurus, dopiero odkryte później i lepiej zachowane czaszki ” przyniosły” obraz konstrukcji zbliżony do czaszki Dromaeosaurusa , jednak nie była ona tak solidna w budowie jak u wcześniej wymienionego. Podniebienie było mocno zaokrąglone, co pozwoliło na zwężenie pyska, co z kolei pozwoliło na ulepszoną wersję wizji stereoskopowej co oznacza, że Deinonychus miał bardzo dobrą percepcję głębokości co pozwalało mu na doskonała ocenę dystansu między nim samym a ofiarą.
Kości jarzmowe były szerokie co sugeruje, że Deinonychus pomimo swoich niewielkich rozmiarów gryzł z stosunkowo wielką siłą (istnieje przypuszczenie, iż największe osobniki były zdolne do przegryzania kości). Czaszka dodatkowo miała bardzo duże fenestry, w tym dwa w dolnej szczęce, które znajdowałby się pod orbitalną fenestrą, gdy pysk były zamknięty.
Ramiona Deinonychusa były długie i stanowiły ponad połowę długości nóg i zakończone były trzema pazurami, badania Carpentera (2002) wykazały, że ramiona Deinonychusa służyły najprawdopodobniej do chwytania ofiar. Ogon zbudowany był podobnie jak u innych członków grupy, z każdym ścięgnem nakładającym się na kilka kręgów powodując, że był on niezwykle sztywny (mógł poruszać się tylko u podstawy). Ogon pełnił swoistą funkcję stabilizatora podczas szybkiego biegu i coś na wzór steru, podczas gdy zwierzę dokonywało gwałtownych zwrotów.
Wielu paleontologów (choć nie wszyscy) uważa, że Deinonychus polował stadnie dowody na to pochodzą z częstych szczątków Deinonychusa znalezionych wokół skamielin Tenontozaurów. Tenontosaurus jest znacznie większy niż Deinonychus, a pojedyncze zwierzę miałoby ogromne trudności z upolowaniem dorosłego Tenontozaurusa, co doprowadziło do ów teorii.
Choć według niektórych naukowców kończyny przednie Deinonychusa były idealnie dostosowane do chwytania zdobyczy to jednak ” śmiertelny arsenał ”stanowił sierpowaty pazur (ok. 12 cm) długości używany do kłucia/dźgania być może i do wbijania go w newralgiczne miejsca na ciele ofiar (np. szyją), jego funkcja mogła opierać się także na celach obronnych przed innymi gatunkami oraz walki o dominację oraz terytorium.
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Nasutoceratops titusi Następny artykuł:
Zuul crurivastator




































Jak zwykle super!
Tak jak wspomniał MAX kolejny świetny artykuł Krzychu btw. uwielbiam twoje opracowania na temat dinozaurów.
Krzysztof, może kiedyś opiszesz Patagotitan mayorum – najnowszego gigantycznego tytanozaura.
Wstępny opis na temat tego tytanozaura już jest umieszczony na DA przez redakcje w artykule ” Najdłuższe zauropody – top 10”. Ja z kolei zapraszam Ciebie oraz innych czytelników na artykuł o dotyczący najlepiej zachowanego dinozaura w historii, który również podobnie jak Patagotitan został niedawno ” ukazany światu ” – Borealopelta markmitchelli, który właśnie przed chwilą zamieściłem.
Znalazłem – Patagotitan mayorum został wstępnie opisany w artykule http://dinoanimals.pl/dinozaury/odkryto-najwiekszego-dinozaura-w-historii/, tuż po odkryciu, kiedy jeszcze nie był nazwany.
Zaraz zabieram się za lekturę prehistorycznej Mona Lizy 🙂
A skąd te dane, że niby Achillo i Austro takie lekkie były?… Hartman wycenił je na 350 i 500 kg, a Ty sporo niższe zakresy podajesz – podobnie zresztą jeśli o Dakota chodzi, wprawdzie Hartman chyba nigdy go nie szacował (a przynajmniej nic mi o tym nie wiadomo) ale porównując go z innymi przedstawicielami tej rodziny to przy takiej długości i wzroście to na pewno w okolicach 0.5 t musiał ważyć, a nie ledwie połowę tego.
P.S. OK, co do Austroraptora to już doczytałam – szacunki są z 2010 r. i pochodzą od Paula, podtrzymuję natomiast pytanie o pochodzenie tych niskich szacunków masy Dakotaraptora i Achillobatora.
Wasze artykuły są świetne,niestety od jakiegoś czasu ich nie czytałem.Czas nadrobić zaległości….Polecam was wszystkim,którzy jeszcze waszych stron nie przeczytali.
Zróbcie coś np.o yi qi,sordes pilosus,opistocelikaudi.Pozdrawiam.
Krzysztof nie opisywał wprawdzie jeszcze jak widzę Yi Qi, ale wejdź sobie na blog Mać Animal World, tak Yi Qi jest opisany.
Dziękuje za słowa uznania i zapraszam do zapoznania się z artykułami, które stworzę w przyszłości.