
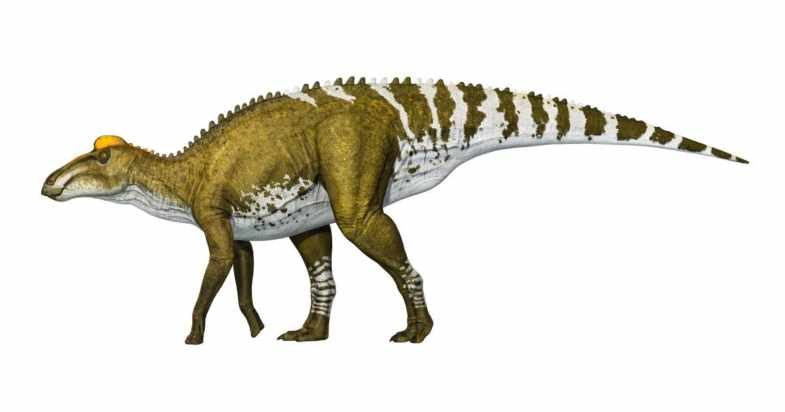
Edmontosaurus regalis.
Edmontosaurus był roślinożernym dinozaurem z rodziny hadrozaurów z podrodziny hadrozaurów płaskogłowych i największym ich przedstawicielem.
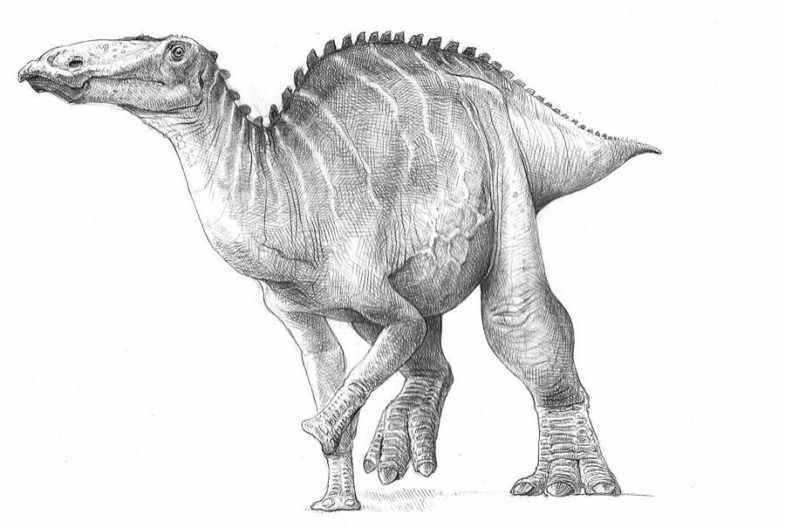
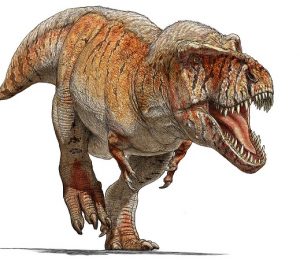
Szkic przedstawiający potencjalny wygląd Edmontosaurus annectens za życia.
Systematyka:
Domena: eukarionty – Królestwo: zwierzęta – Typ: strunowce – Podtyp: kręgowce
Gromada: zauropsydy – Podgromada: diapsydy – Nadrząd: dinozaury
Rząd: dinozaury ptasiomiednicze – Podrząd: cerapody – Infradząd: ornitopody
Nadrodzina: hadrozauroidy – Rodzina: hadrozaury – Podrodzina: hadrozaury płaskogłowe
Rodzaj: Edmontosaurus – Gatunki: Edmontosaurus annectens / regalis/saskatchewanensis
Czas i miejsce występowania:
• Edmontosaurus annectens: Stany Zjednoczone– Wyoming (formacja Lance), Montana (formacja Hell Creek), Dakota Południowa (formacje Lance i Hell Creek). Kolorado (formacja Laramie), Kanada – Alberta (formacja Scollard), Saskatchewan (formacja Frenchman) ok. 68 –66 mln lat temu późna kreda (późny mastrycht).• Edmontosaurus regalis: Kanada – Alberta (formacja Horseshoe Canyon – ogniwo Horsethief, formacja St Mary River, formacja Wapiti) ok. 72,6 – 71 mln lat temu późna kreda ( kampan/mastrycht).• Edmontosaurus sasktchewanensis:
Kanada – późny mastrycht
Etymologia:
Nazwa Edmontosaurus oznacza jaszczura z Edmonty i przypuszczalnie pochodzi od formacji (dziś grupy) Edmonton.
Szacunki długości i masy:
• Edmontosaurus annectens MOR 1142 – ”X rex” (największy przedstawiciel rodzaju):
W 2011 roku Horner i Woodward na Hadrosaur Symposium określili długość tego osobnika w przedziale 15 – 16,1 (sama długość ogona określana jest na 7,6 metra), szacunek masy podawany przez wcześniej wymienionych to minimalnie 14000, maksymalny to aż 40000 kilogramów (gdzie najbardziej prawidłowym przedziałem była by masa od 14000 do 24000 kilogramów).
•Brett-Surman (1997) 12 – 13 metrów, Morris (1970) 12,2 metra •Horner i Woodward (2011) 12 – 13 metrów dla okazu MOR 1609 (Becky’s Giant), masa ciała określana jest na ponad 10000 kilogramów
• Paula online (2010), Benson i współpracownicy (2014) 10 metrów długości, masa w przedziale 5000 – 10000 kilogramów dla osobnika MOR 003
• Campione i Evansa (2011) 8,8 metra dla okazu AMNH 5730, masa 3200 – 6600 kilogramów Paula online.
• Edmontosaurus regalis:
12,2 metra dla holotypu według Lambe (1920) oraz 9 – 12 metrów według Lull i Wight (1942), masa w przedziale 7000 – 8000 kilogramów
• Edmontosaurus saskatchewanensis:
Gatunek ten jest najmniejszym przedstawicielem rodzaju szacowanym na 7 metrów długości, choć prawdopodobnie osobnik był jeszcze w fazie wzrostu, masa ciała 3000 – 4000 kilogramów

Model przedstawiający potencjalny wygląd Edmontosaurus annectens za życia.
Odkrycie:
Dwa pierwsze okazy Edmontosaurus (USNM 2414 / YPM 2182 – paratyp) zostały znalezione w 1891 roku przez Johna Bella Hatchera w okręgu Niobrara (wówczas część Converse County) w Wyoming, który datuje się na okres późnej kredy (mastrycht).
Klasyfikacja:
Edmontosaurus należy do jednej z dwóch podrodzin hadrozaurydów – Hadrosaurinae, ostatnio przemianowanych na Saurolophinae, bo Hadrosaurus okazał się być bardziej bazalny (Prieto-Márquez, 2011). Pozycja filogenetyczna Edmontosaurus jest niepewna. Wg wielu analiz jest on dość bazalnym przedstawicielem Saurolophinae a według innych jest zaawansowany i bliski kladowi Prosaurolophus+Saurolophus oraz azjatyckim rodzajom Kerberosaurus i Kundurosaurus. Najbliższym krewnym Edmontosaurus może być Shantungosaurus (Gates i współpracownicy 2011; Prieto-Márquez, 2013). Jak wynika z analizy Prieto-Márqueza (2010) i jej modyfikacji, mimo późnego okresu występowania (Edmontosaurus to ostatni północnoamerykański dinozaur kaczodzioby), Edmontosaurus jest dość bazalnym przedstawicielem Saurolophinae, którego linia oddzieliła się od innych zaurolofinów na początku kampanu albo nawet wcześniej (ta luka w zapisie kopalnym, największa wśród północnoamerykańskich hadrozaurydów, wskazuje, że przodkowie Edmontosaurus wciąż czekają na odkrycie). Jedna z wielu jego analiz wykazała jednak, że hadrozaurydy powszechnie zaliczane do Hadrosaurinae (Saurolophinae) nie tworzą kładu, ale grad wiodący ku grzebieniastym lambeozaurynom. W tym wypadku Edmontosaurus jest bliski przedstawicielom Lambeosaurinae. Bazalną pozycję wśród Saurolophinae wskazują też inne analizy (Prieto-Marquez (2005) Prieto-Marquez i Wagne (2009) Dalla Vecchia (2009) Xing i współpracownicy (2012). Wiele badań wskazuje jednak, że Edmontosaurus jest zaawansowany i (czasem razem ze Shantungosaurus – Gates i współpracownicy (2011) jest taksonem siostrzanym kładu Prosaurolophus+Saurolophus (Weishampel i współpracownicy (1993), Gates i Sampson (2007) Prieto-Marquez i Salinas (2010), zawierającego prawdopodobnie Kerberosaurus (Bolotsky i Godefroit (2004) Godefroit i współpracownicy (2008) Bell (2011) Gates i współpracownicy (2011) Xing i współpracownicy (2012) Xing i współpracownicy (2014). Natomiast analiza Godefroita i współpracowników (2012) wskazuje, że Kerberosaurus i Kundurosaurus należą do kładu z Edmontosaurus i wraz z nim są taksonem siostrzanym Prosaurolophus+Saurolophus; podobnie analiza Suesa i Averianova (2009) (niezawierająca nieopisanego wtedy Kundurosaurus). Analiza Hornera i współpracowników (2004) wykazała natomiast, że Edmontosaurus jest taksonem siostrzanym Maiasaura i Brachylophosaurus.
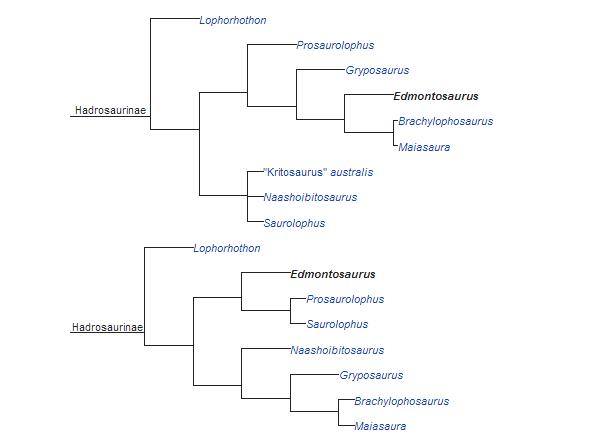
Kładogramy ukazujące pozycję systematyczną Edmontosaurus (u góry autorstwa Hornera –2004, na dole autorstwa Gatesa i Sampsona – 2007).
Czaszka:
Czaszka w pełni dorosłego osobnika Edmontosaurus oscylowała w granicach 120 – 150 cm i miała z grubsza trójkątny profil. Widziana z góry przednia i tylna część czaszki ulegały rozszerzeniu, a szeroki przód tworzył swoisty kształt kaczego dziobu. Dziób ten był bezzębny, gdzie jego górna i dolna część zawierały materiał keratynowy (pozostałości szczątkowego keratynowego górnego dzioba znane są z ”mumii” Edmontosaurus przechowywanej w Muzeum Senckenberga). W okazie z wcześniej wspomnianego muzeum zachowana bezkostna część dzioba rozciąga się co najmniej 8 cm poza kość wystając pionowo w dół. Otwory nosowe Edmontosaurus były wydłużone i osadzone w głębokich zagłębieniach otoczonych wyraźnymi obrzeżami kostnymi. W przynajmniej jednym okazie z Naturmuseum Senckenberg w oczodołach zachowała się bardzo rzadko spotykany pierścień twardówkowy. Inna rzadko spotykaną kością jest strzemiączko (gadzia kość słuchowa) zaobserwowaną również w okazach prezentowanych w wyżej wymienionym muzeum. Zęby składały się z sześciu rodzajów tkanek i rosły w kolumnach (51 do 53 kolumn w górnej szczęce i 48 do 49 w dolnej dla E.regalis i 52 kolumny w górnej szczęce i 44 w dolnej dla E. annectens ).
W 2011 roku Campione i Evans zanotowali dane z badań nad wszystkimi znanymi czaszkami Edmontosaurus datowanych na Mastrycht i Kampan i wykorzystali je do wykreślenia wykresu morfometrycznego porównującego zmienne cechy czaszki wraz z ich rozrostem. Ich wyniki pokazały, że w obu uznanych gatunkach Edmontosaurus wiele cech używanych wcześniej do klasyfikowania dodatkowych gatunków lub rodzajów było bezpośrednio zestawiane z rozmiarem czaszki. Campione i Evans interpretowali te wyniki jako silnie sugerujące, że kształt czaszek Edmontosaurus zmieniał się dramatycznie wraz z ich wzrostem. Doprowadziło to do kilku oczywistych błędów w klasyfikacji dokonywanych w przeszłości np. gatunek Thespesius edmontoni uważany wcześniej za synonim E. annectens ze względu na jego mały rozmiar i kształt czaszki jest prawdopodobnie młodocianym okazem E. regalis.

Kompletne czaszki poszczególnych gatunków rodzaju Edmontosaurus.
Mózg:
Mózg Edmontosaurus został opisany w kilku artykułach i streszczeniach poprzez użycie odlewu wnętrza czaszki, w której znajdował się mózg E. annectens oraz E. regalis. Mózg nie był szczególnie duży jak na zwierzę wielkości Edmontosaurus, przestrzeń, w której się mieścił stanowiła tylko około ćwierć długości czaszki, a różne odlewy dawały pomiar od 374 do 450 mililitrów co oznacza, że mózg zajmować jedynie ok. 50% przestrzeni wnętrza czaszki resztę przestrzeni zajmowała opona twarda (jedna z opon mózgowo-rdzeniowych) otaczająca mózg.
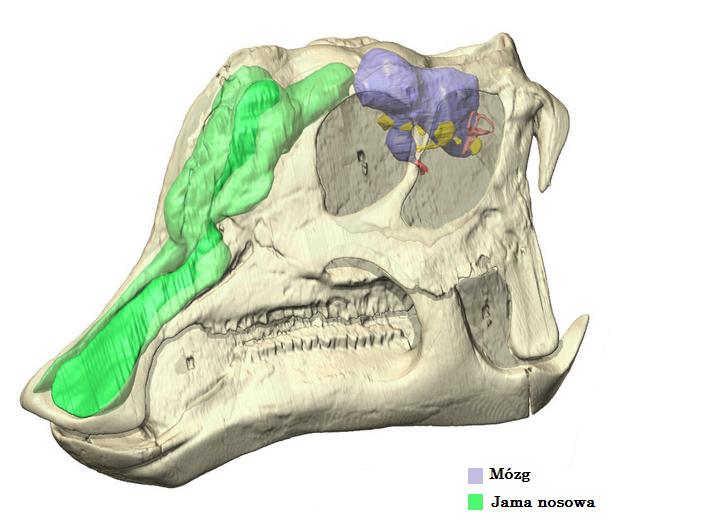
Art przedstawiający wielkość mózgu Edmontosaurus.
Budowa anatomiczna szkieletu:
Liczba kręgów różni się między okazami i tak Edmontosaurus regalis miał 13 kręgów szyjnych, 18 kręgów kręgosłupa, 9 kręgów biodrowych i 85 kręgów ogonowych (inne mają od 50 do 70). Przednia część pleców była zakrzywiona w kierunku ziemi, z szyją zgiętą do góry, a reszta grzbietu i ogona trzymana poziomo. Większość grzbietu i ogona była ”wyłożona” skostniałymi ścięgnami ułożonymi w kratownicę wzdłuż kręgów nerwowych kręgosłupa (skostniałe ścięgna są interpretowane jako wzmacniające kręgosłup przeciw naprężeniom grawitacyjnym). Ramiona/ łopatki były w kształcie podobne do ostrza trzymanymi w pozycji mniej więcej równoległej do kręgosłupa. Biodra składały się z trzech elementów: kości biodrowej znajdującej się po wyżej stawu skokowego, kości kulszowej poniżej i kości łonowej z przodu. Ramiona miały duży obszar obojczykowy, gdzie ”przymocowane” były mięśnie, kość łokciowa i promieniowa były ”szczupłe” w budowie. Ramiona i przedramiona miały podobną długość, nadgarstek był prosty i zbudowany był z dwóch małych kości, a każda z ”rąk” miała cztery palce. Drugi, trzeci i czwarty palce były w przybliżeniu tej samej długości i łączyły się w mięsistym pokryciu. Mały palec oddzielił się od pozostałych trzech i był znacznie krótszy. Kość udowa była solidna i prosta, z widocznym kołnierzem około połowy tylnej strony. Długi ogon pomagał w utrzymaniu balansu ciała.
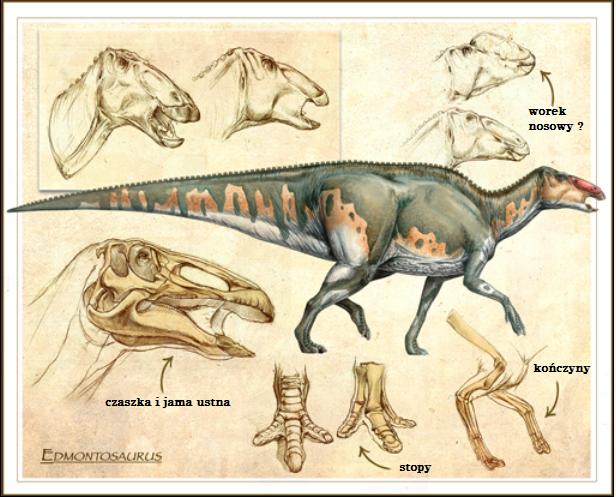
Art. przedstawiający budowę anatomiczną wybranych fragmentów ciała Edmontosaurus.
Mechanizm żuchwy:
Edmontosaurus był lądowym roślinożercą, który posiadał setki zębów (od 1500 – 2000) z których tylko względna ilość była używana w dowolnym momencie żerowania (zasięg żerowania zwierzęcia wynosił ok. 4 metrów od poziomu podłoża wzwyż). Swojego szerokiego dzioba używał do zrywania z gałęzi liści i pędów. Edmontosaurus jak i inne dinozaury ptasiomiednicze posiadał muskularne policzki, których funkcją było zatrzymywanie jedzenia w jamie ustnej. W latach 1960 – 1970 dominującą interpretacją hadrozaurów był ich nadwodny styl życia i pożywianie się roślinnością wodną – przykładem tego jest interpretacja Williama Morrisa z 1970 roku. Morris zaproponował on sugestię, w której zwierzę miało dietę podobną do współczesnych kaczek, filtrując roślinność i morskie bezkręgowce przez bruzdy w kształcie litery V wzdłuż wewnętrznej powierzchni górnego dzioba. Interpretacja ta została jednak odrzucona, ponieważ bruzdy przypominają te jakie występują u współczesnych trawożernych żółwi niż elastyczne struktury obserwowanych u ptaków, które filtrują wodę w pozyskaniu pożywienia. Od połowy lat osiemdziesiątych do pierwszej dekady 2000 roku dominującą interpretacją sposobu, w jaki hadrozaury przetwarzały żywność, była zgodna z modelem przedstawionym w 1984 roku przez Davida B. Weishampela. Stwierdził on, że struktura czaszki pozwalała na ruch między kościami, co powodowało ruchy żuchwy w przód i w tył, oraz wygięcie na zewnątrz zębów kości szczęki, gdy usta były zamknięte. Zęby górnej szczęki ocierały by się o dolne niczym tarki a materiał roślinny był uwięziony między nim, taki ruch byłby równoległy do efektów żucia u ssaków, chociaż osiągałby efekty w zupełnie inny sposób.
Prace na początku 2000 roku zakwestionowały model Weishampela. W badaniu opublikowanym w 2008 r. przez Caseya Hollidaya i Lawrence’a Witmera okazało się, że ornitopody, takie jak Edmontosaurus, nie posiadały typów stawów czaszkowych obserwowanych u tych współczesnych zwierząt, o których wiadomo, że mają czaszki kinetyczne (czaszki umożliwiające ruch pomiędzy ich składowymi kośćmi), takie jak łuskonośne (rząd w gromadzie gadów) i ptaki. Zaproponowali oni, że stawy, które zostały zinterpretowane jako pozwalające na ruch w czaszkach dinozaurów, były w istocie chrząstkowymi strefami wzrostu. Ważnym dowodem dla modelu Weishampela jest ukierunkowanie zadrapań na zębach, pokazująca kierunek działania szczęki i ruchu kości obu połówek żuchwy. Vincent Williams i współpracownicy (2009) w swojej opublikowanej pracy ” hadrosaurid tooth microwear” przedstawili cztery klasy zadrapań na zębach Edmontosaurus, najczęściej spotykana klasa była interpretowana jako wynik skośnego ruchu, a nie prostego ruchu w górę lub w dół, co jest zgodne z modelem Weishampla (uważa się, że ten ruch był podstawowym ruchem mielenia żywności). Dwie kolejne klasy zarysowań zostały zinterpretowane jako będące wynikiem ruchów szczęk w przód lub w tył. Czwarta klasa była zmienna i prawdopodobnie wynikała z otwarcia szczęk, gdzie kombinacja ruchów była bardziej złożona, niż przewidywano.
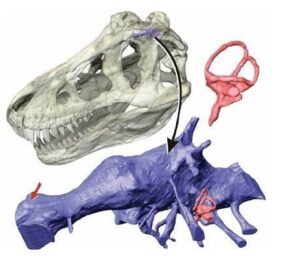
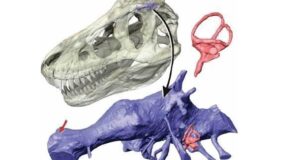
Weishampel opracował swój model za pomocą symulacji komputerowej. Natalia Rybczynski wraz z współpracownikami zaktualizowała tę pracę o wiele bardziej wyrafinowanym trójwymiarowym modelem animacji, skanując czaszkę E. regalis za pomocą laserów. Byli oni w stanie powtórzyć proponowany ruch w swoim modelu chociaż odkryli, że potrzebne są dodatkowe ruchy wtórne między innymi kościami, z maksymalnym rozdziałem od 1,3 do 1,4 centymetra między niektórymi kośćmi podczas żucia.
W 2012 roku badania opublikowane przez Robina Cuthbertsona i współpracowników wykazały, że ruchy wymagane w modelu Weishampela są mało prawdopodobne, i faworyzowały model, w którym ruchy żuchwy wywoływały ostry ruch. Połączenie górnej i dolnej szczęki umożliwiały ruch przednio – tylny wraz ze zwykłym obrotem, a przednie stawienie dwóch połówek żuchwy również umożliwiało ruch (w połączeniu dwie połowy dolnej szczęki mogły poruszać się lekko w przód i w tył, a także obracać się nieznacznie wzdłuż ich długich osi).
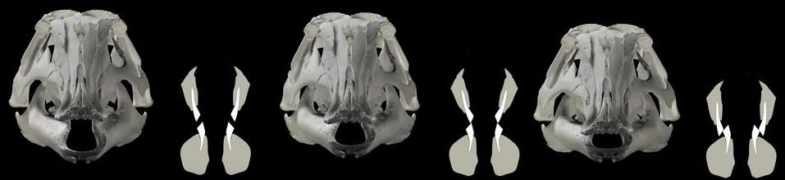
Schemat mechanizmu żuchwy Edmontosaurus.
Badania izotopowe/dieta:
Okaz ”mumii”, która wchodzi w skład kolekcji należącej do Senckenberg Museum posiadał w tuszy zawartość jelit. Charles H. Sternberg zgłosił obecność karbonizowanej treści jelit w okazie. Pokarm roślinny znajdujący się w tuszy okazu znajdującego się w Senckenberg Museum został opisany a stanowiły go igły drzewa Cunninghamites elegans, gałązki innych drzew iglastych i liściastych oraz liczną ilość małych nasion i owoców.
Williams i współpracownicy badając strukturę zadrapań struktury mikroukładu zębów sugerowali. że Edmontosaurus był zwierzęciem pasącym się roślinnością bogatą w zawartość krzemionki takimi jak skrzypy i paprocie.
Dieta i fizjologia Edmontosaurus została zbadana za pomocą stabilnych izotopów węgla i tlenu zarejestrowanych w szkliwie zębów. Podczas żerowania, picia oraz oddychania zwierzę pobierało węgiel i tlen, który został wkomponowany do kości. Izotopy tych dwóch pierwiastków określają różne czynniki wewnętrzne i zewnętrzne, takie jak rodzaj spożywanych roślin, fizjologia zwierząt, zasolenie i klimat. Jeśli proporcje izotopów w skamieniałościach nie zostały by zmienione przez fosylizację i późniejsze zmiany, mogą być one badane w celu uzyskania informacji o pierwotnych czynnikach, gdzie zwierzęta stałocieplne miały by pewne składy izotopowe porównywalne do ich otoczenia, a zwierzęta, które żywią się pewnymi typami roślinności lub stosują określone procesy trawienne będą miały wyraźne kompozycje izotopowe.
Kathryn Thomas i Sandra Carlson w swoim badaniu z 2004 roku wykorzystały górne szczęki trzech osobników (młodego, nastoletniego i dorosłego), gdzie żeby wszystkich trzech osobników zdawały się pokazywać zmiany stosunków izotopów tlenu, które mogą odpowiadać okresom ciepłym / suchym i chłodnym / mokrym. Thomas i Carlson rozważały możliwość migracji zwierząt w badaniu, ale bardziej faworyzowały lokalne wahania sezonowe, ponieważ migracja prawdopodobnie prowadziłaby do homogenizacji proporcji, ponieważ wiele zwierząt migruje, aby utrzymać się w określonych zakresach temperatur lub w pobliżu konkretnych źródeł żywności. Edmontosaurus wykazywał również wzbogacone wartości izotopów węgla, które dla współczesnych ssaków można by interpretować jako mieszaną dietę roślin C3 (większość roślin) i roślin C4 (traw); jednak rośliny C4 były niezwykle rzadkie w późnej kredzie, jeśli w ogóle były obecne. Thomas i Carlson sugerują również, że w skład diety Edmontosaurus mogły wchodzić również rośliny zawierające chlorek sodu, które występowały w obszarach rejonów przybrzeżnych (Western Interior Seaway).

Mumia Edmontosaurus annectens – Naturmuseum Senckenberg (Frankfurt, Niemcy).
Skóra:
Znaleziono wiele okazów Edmontosaurus annectens z dobrze zachowanymi fragmentami skóry. Kilka z nich zyskało rozgłos w śród nich chyba jednym z najsłynniejszych jest ”Trachodon mummy”odkryta w początku XX wieku, której nadano ksywę ”Dakota”, która zawierała resztkowe związki organiczne ze skóry. Z powodu tych odkryć, skalowanie E.annectens jest znane dla większości obszarów ciała. Odciski skóry są mniej znane z okazu E.regalis, ale zbadano kilka dobrze zachowanych okazów w tym jeden, który zachował tkanki miękkie grzebienia na łubie.
W odniesieniu do wielkości ciała zwierzęcia, skóra wydaje się wydaje się cienka i delikatna. Jako typowa dla nieopierzonych dinozaurów skóra składała się z niepokrywających się łusek/guzków, gdzie można było wyróżnić ich dwa rodzaje: małe (o średnicy od 1 do 3 mm) zaokrąglone bulwy gruntowe, które były równomiernie rozmieszczone oraz większe (mniej więcej 5 mm średnicy) guzki brukowate mają kształt pięciokąta, które były nieco uniesione i rozmieszczone w nieregularnych skupiskach. Klastry składały się od dwudziestu do kilkuset pojedynczych guzków brukowych otoczonych przez pośrednie guzki, które pośredniczyły w rozmiarze i kształcie między guzkami brukowatymi. W okolicy klatki piersiowej i brzucha klastry były małe, owalne i ułożone w nieregularnych podłużnych liniach, stawały się one większe w okolicach boków tułowia, gdzie osiągały średnicę 5 do 10 mm a ich kształt stawał się bardziej nieregularny. Największe znajdowały się nad miednicą (50 mm), gdzie tworzyły największe skupiska, przypuszczalnie gromady o podobnej wielkości były obecne wzdłuż grzbietu zwierzęcia.

Materiał skalny okazu Edmontosaurus regalis z zachowanym fragmentem skórnym.
Lokomocja:
Podobnie jak inne hadrozaury, Edmontosaurus uważany był za zwierzę poruszające się głównie na czterech kończynach, ale w razie potrzeby mógł przyjąć postawę dwunożną. Prawdopodobnie na czworakach poruszał się powoli, przestawiając się na używanie tylnych kończyn, gdy chciał poruszać się szybciej (np. ucieczka przed drapieżnikiem). Badania przeprowadzone w 2007 roku za pomocą modelowania komputerowego na młodocianym osobniku o wadzę około 715 kilogramów sugerowały, że mógł on poruszać się z dużą prędkością, która mogła dochodzić nawet do 45 km/h.

Edmontosaurus mógł poruszać się zarówno na czterech jak i dwóch kończynach.
Interakcja z drapieżnikami:
Przedział czasowy i zasięg geograficzny Edmontosaurus pokrywały się z Tyrannosaurus rex, a dorosły osobnik E. annectens prezentowany w Denver Museum of Nature and Science pokazuje dowód potencjalnego ataku Tyrannosaurus w postaci ugryzienia w ogon. Odwracając się od strony bioder kręgi od XIII do XVII mają uszkodzone grzbiety odpowiadające atakowi z prawej tylnej części zwierzęcia. Jeden fragment kręgu ogonowego jest wyłamany pozostałe są zagięte, choć trzy z nich mają wyraźne ślady po przebiciu zębów. Wierzch ogona miał wysokość co najmniej 2,9 m, a jedynym gatunkiem teropoda znanym z tej samej formacji skalnej, który były wystarczając wysoki, by wywołać taki atak, był T. rex. Kości zaatakowanego osobnika były częściowo wyleczone, ale Edmontosaurus zdechł zanim ślady uszkodzeń zostały całkowicie zatarte, uszkodzenie wykazywało również oznaki infekcji kości. Kenneth Carpenter, który badał okaz, zauważył, że zdaje się, że w lewym biodrze wyleczone zostało złamanie poprzedzające atak, ponieważ było ono w pełni wyleczone, zasugerował, że Edmontosaurus był celem, ponieważ mógł kuleć z powodu wcześniejszych obrażeń.
Inny okaz E. annectens o potencjalnej długości 7,6 metra z Dakoty Południowej, wykazuje ślady ugryzień, które przypisuje się małym teropodom w okolicach dolnej szczęki, które zostały częściowo wyleczone. Michael Triebold sugeruje scenariusz, w którym zwierzę zostało zaatakowane przez niewielkie teropody, które atakowały gardło Edmontosaurus przeżył atak, ale uległ obrażeniom między innymi w wcześniej wspomnianej okolicy.

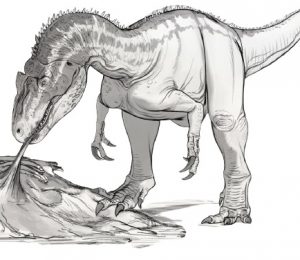
Szkic przedstawiający atak grupowy Tyrannosaurus rex na Edmontosaurus annectens.
Nowotwory:
W 2003 roku w kościach Edmontosaurus opisano szereg nowotworów między innymi: naczyniaki krwionośne, włókniaki desmoplatyczne, kostniaki zarodkowe oraz raka przerzutowego. Rothschild i współpracownicy przebadali kręgosłup dinozaura pod kątem nowotworów za pomocą tomografii komputerowej i badania fluoroskopowego. Kilka innych hadrosaurów, w tym Brachylophosaurus, Gilmoreosaurus i Bactrosaurus, również wykazywało skłonności do powstawania w ich organizmie zmian nowotworowych. Chociaż zbadano w ten sposób ponad 10000 skamielin, guzy ograniczono do Edmontosaurus i blisko spokrewnionych rodzajów. Guzy mogły być spowodowane czynnikami środowiskowymi lub skłonnością genetyczną. Osteochondroza, czyli powierzchowne dołki w kościach w miejscach, w których wyartykułowane są kości, jest również znana w Edmontosaurus, stwierdzono, że to stan, wynikający z braku wymiany chrząstki przez kości podczas wzrostu, występujący w 2,2% z 224 kości Edmontosaurus. Przyczyna tego stanu nie jest znana, sugerowano predyspozycje genetyczne, traumę, intensywność karmienia, zmiany w dopływie krwi, nadmiar hormonów tarczycy i niedobory różnych czynników wzrostu.
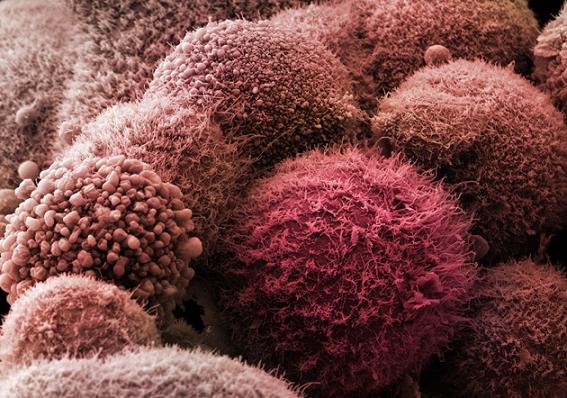
Edmontosaurus był rodzajem dinozaura, który prawdopodobnie w wyniku skłonności genetycznych był narażony na występowanie nowotworów.
Paleoekologia/paleobiologia:
Edmontosaurus był rodzajem rodzajem rozpowszechnionym zarówno w czasie jak i przestrzeni. Jednostki skalne, z których jest znany można podzielić na dwie grupy wiekowe: starsze – formacje: Horseshoe Canyon i St. Mary River oraz młodsze – formacje: Hell Creek, Lance, Frenchman. Okres czasu, który obejmował formację Horseshoe Canyon jest również znany jako Edmontonian, a okres czasu obejmowany przez młodsze jednostki znany jest jako Lancian (interwały czasowe Edmontonian i Lancian miały odrębne dinozaury). Edmontonian jest określany jako epoka, w której po raz pierwszy pojawił się gatunek Edmontosaurus regalis (chociaż czasami opisywano go jako pochodzący wyłącznie z wczesnego Matstrychtu). Osadzanie rozpoczęło się około 73 milionów lat temu, pod koniec Kampanu i zakończyło się między 68 a 67,6 milionów lat temu. Edmontosaurus regalis jest znany z najniższego z pięciu pokładów formacji Horseshoe Canyon (ale jest nieobecny przynajmniej od drugiego aż do szczytu, a aż trzy czwarte okazów dinozaurów z badlands w pobliżu Drumheller, Alberta może odnosić się do rodzaju). Lancian był ostatnim interwałem czasowym poprzedzającym wielkie wymieranie kredowe.
Formacja Lance, w której występował Edmontosaurus regalis klimat wilgotny/subtropikalny, roślinność stanowiły: drzewa iglaste, liściaste, jesion, dąb, krzewy, paprocie. Oprócz dinozaurów faunę stanowiły: ryby słodkowodne, salamandry, żółwie. Różnorakie jaszczurki, węże, ptaki i małe ssaki. Małe dinozaury nie występowały w tak dużej obfitości jak w Hell Creek, ale Thescelosaurus po raz wydawał się być dość powszechny, Triceratops natomiast wydawał się być nieco mniejszy rozmiarowo co potwierdza wielkość odnajdowanych czaszek z tej formacji. Oprócz dwóch wcześniej wymienionych w skład fauny wchodziły: Hypacrosaurus altispinus, Pachyrhinosaurus canadensis, Anchiceratops ornatus, Edmontonia longiceps, Arrhinoceratops brachyops, Saurolophus osborni, Atrociraptor marshalli, Troodon formosus, Dyoplosaurus acutosquameus, Albertosaurus sarcpohagus.
W formacji Hell Creek Edmontosaurus annectens oprócz wcześniej wspomnianego Triceratops horridus oraz Thescelosaurus neglectus koegzystował z takim dinozaurami jak: Tyrannosaurus rex, Ankylosaurus magniventris, Struthiomimus altus, Ornithomimus velox, Anzu wylei, Pachycephalosaurus wyiomingensis, Stygimoloch spinifer, Dracorex hogwarista, Sphaerotholus buchholtzae, Archeoraptor temertyorum, Laptoceratops gracilis, Torosaurus latus.

Edmontosaurus w towarzystwie ceratopsa: Anchiceratops ornatus, aligatorida: Brachychampsa gilmore oraz żółwia z rodzaju Trionyx.
Materiał kopalny:
Holotyp to USNM 2414 to częściowa górna część czaszki i szkielet. Paratyp to YPM 2182. Przypisane okazy to: AMNH 427: górna część czaszki i mózgoczaszka, AMNH 5046: częściowa czaszka młodego okazu, AMNH 5060: kompletna czaszka i szkielet („mumia”), AMNH 5046: częściowa czaszka (brak pyska) młodego okazu, AMNH 5730 (holotyp E. copei), kompletna czaszka i szkielet, BHI 2169: kompletna dysartykułowana czaszka, CCM (bez nr): częściowa czaszka i kompletny szkielet, CMN 8509 (holotyp E. saskatchewanensis): kompletna czaszka i częściowy szkielet, DMNH 1493, LACM 23502: kompletna czaszka, MOR 003: kompletna czaszka, MOR 1627, NCSM 23119: kompletna czaszka, ROM 57100: kompletna czaszka, SM R4050: kompletna czaszka i szkielet, UCMP 128372: kompletna czaszka, UMMP 20000: kompletna czaszka, USNM 3814: kompletna czaszka i niekompletny szkielet.

Sekwencja ataku Tyrannosaurus rex na Edmontosaurus annectens – Houston Museum of Natural Science (Stany Zjednoczone).
Podobne artykuły
-

‘’Cudzych chwalimy swego nie znamy sami nie wiemy co posiadamy’’
-
”Intelektualiści” ?
-

‘’Hospodar wyspy Hateg’’
Shastasaurus Następny artykuł:
Shantungosaurus giganteus / Zhuchengosaurus maximus / Huaxiaosaurus aigahtens





































Piękny dinozaur i wspaniały artykuł Krzysztof!
Hej Krzysztof,polecałbym zrobić artykuł o kulindadromeusie.
Hehe, JBon, a czemu go sam nie zrobisz :)?
Z tymi nowotworami to kiepska sprawa. Może wszystkie dinozaury w kredzie były podatne na nowotwory i dlatego wymarły?
Dobry artykuł jak zawsze Krzychu, wielki ten Xrex.
Edmontozaury były naprawdę wielkie, dłuższe i cięższe od Tyranozaurów. Czy wiadomo, jak często dorosłe Edmontozaury były atakowane przez drapieżniki?
Witaj Przemku Edmontosaurus zapewne stanowił dość częsty obiekt ataku Tyrannosaurus z tego względu iż był zapewne zwierzyną łowna która nie stanowiła zbyt potencjalnie zbyt dużego ryzyka urazu podczas polowania jak np. w Triceratops czy Ankylosaurus.